Aquaculture
Zoo.667
Fish Physiology

Fish physiology is the scientific study of how the component parts of fish function together in the living fish. It can be contrasted with fish anatomy, which is the study of the form or morphology of fishes. In practice, fish anatomy and physiology complement each other, the former dealing with the structure of a fish, its organs or component parts and how they are put together, such as might be observed on the dissecting table or under the microscope, and the later dealing with how those components function together in the living fish.
Aquatic respiration
Aquatic respiration is the process whereby an aquatic animal obtains oxygen from water.
Fish
In most fish respiration takes place through gills. Lungfish, however, possess one or two lungs. The labyrinth fish have developed a special organ that allows them to take advantage of the oxygen of the air, but is not a true lung. Fish use the process known as countercurrent flow, in which water and blood flow in opposite directions across the gills, maximizing the diffusion of oxygen.
Molluscs
Molluscs generally possess gills that allow exchange of oxygen from an aqueous environment into the circulatory system. These animals also possess a heart that pumps blood which contains hemocyaninine as its oxygen-capturing molecule. Therefore, this respiratory system is similar to that of vertebrate fish. The respiratory system of gastropods can include either gills or a lung.
Arthropods
Aquatic arthropods generally possess some form of gills in which gas exchange takes place by diffusing through the exoskeleton. Others may breathe atmospheric air while remaining submerged, via breathing tubes. In all cases, oxygen is provided from air trapped by hairs around the animals body.
Aquatic reptiles
All aquatic reptiles breathe air into lungs. The anatomical structure of the lungs is less complex in reptiles than in mammals, with reptiles lacking the very extensive airway tree structure found in mammalian lungs. Gas exchange in reptiles still occurs in alveoli however, reptiles do not possess a diaphragm. Thus, breathing occurs via a change in the volume of the body cavity which is controlled by contraction of intercostal muscles in all reptiles except turtles. In turtles, contraction of specific pairs of flank muscles governs inspiration or expiration.
Amphibians
Both the lungs and the skin serve as respiratory organs in amphibians. The skin of these animals is highly vascularized and moist, with moisture maintained via secretion of mucus from specialized cells. While the lungs are of primary importance to breathing control, the skin's unique properties aid rapid gas exchange when amphibians are submerged in oxygen-rich water.
Aquatic birds
The respiratory system of birds differs significantly from that found in mammals, containing unique anatomical features such as air sacs. The lungs of birds also do not have the capacity to inflate as birds lack a diaphragm and a pleural cavity. Gas exchange in birds occurs between air capillaries and blood capillaries, rather than in alveoli.
Fish Respiration
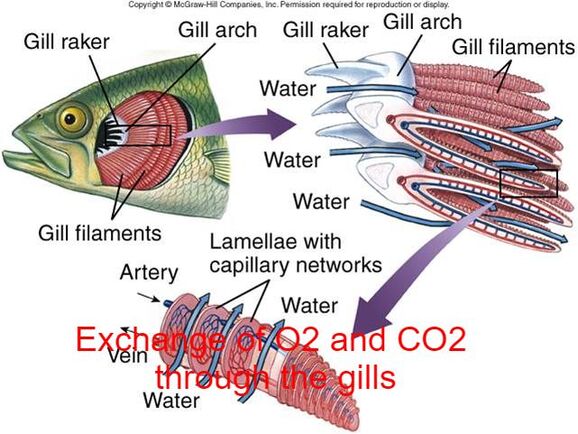
Most fish exchange gases using gills on either side of the pharynx (throat). Gills are tissues which consist of threadlike structures called filaments. These filaments have many functions and "are involved in ion and water transfer as well as oxygen, carbon dioxide, acid and ammonia exchange.
The gills consist of a series of gill arches which have gill filaments on one side and gill rakers on the other. The gill filaments which are used for respiration.
Aquatic respiration
Aquatic respiration is the process whereby an aquatic animal obtains oxygen from water.
Fish
In most fish respiration takes place through gills. Lungfish, however, possess one or two lungs. The labyrinth fish have developed a special organ that allows them to take advantage of the oxygen of the air, but is not a true lung. Fish use the process known as countercurrent flow, in which water and blood flow in opposite directions across the gills, maximizing the diffusion of oxygen.
Molluscs
Molluscs generally possess gills that allow exchange of oxygen from an aqueous environment into the circulatory system. These animals also possess a heart that pumps blood which contains hemocyaninine as its oxygen-capturing molecule. Therefore, this respiratory system is similar to that of vertebrate fish. The respiratory system of gastropods can include either gills or a lung.
Arthropods
Aquatic arthropods generally possess some form of gills in which gas exchange takes place by diffusing through the exoskeleton. Others may breathe atmospheric air while remaining submerged, via breathing tubes. In all cases, oxygen is provided from air trapped by hairs around the animals body.
Aquatic reptiles
All aquatic reptiles breathe air into lungs. The anatomical structure of the lungs is less complex in reptiles than in mammals, with reptiles lacking the very extensive airway tree structure found in mammalian lungs. Gas exchange in reptiles still occurs in alveoli however, reptiles do not possess a diaphragm. Thus, breathing occurs via a change in the volume of the body cavity which is controlled by contraction of intercostal muscles in all reptiles except turtles. In turtles, contraction of specific pairs of flank muscles governs inspiration or expiration.
Amphibians
Both the lungs and the skin serve as respiratory organs in amphibians. The skin of these animals is highly vascularized and moist, with moisture maintained via secretion of mucus from specialized cells. While the lungs are of primary importance to breathing control, the skin's unique properties aid rapid gas exchange when amphibians are submerged in oxygen-rich water.
Aquatic birds
The respiratory system of birds differs significantly from that found in mammals, containing unique anatomical features such as air sacs. The lungs of birds also do not have the capacity to inflate as birds lack a diaphragm and a pleural cavity. Gas exchange in birds occurs between air capillaries and blood capillaries, rather than in alveoli.
Fish Respiration
Most fish exchange gases using gills on either side of the pharynx (throat). Gills are tissues which consist of threadlike structures called filaments. These filaments have many functions and "are involved in ion and water transfer as well as oxygen, carbon dioxide, acid and ammonia exchange.
The gills consist of a series of gill arches which have gill filaments on one side and gill rakers on the other. The gill filaments which are used for respiration.
Air typically contains 280 mg/l of oxygen while fresh water typically only contains 5 - 10 mg/l.
Most fishes will be satisfied with oxygen levels of 5 or 6 mg/l of free oxygen in their environment but if this level drops the fish will be seen gasping at the surface in an attempt to find the most oxygen.
Each filament contains a capillary network that provides a large surface area for exchanging oxygen and carbon dioxide. Fish exchange gases by pulling oxygen-rich water through their mouths and pumping it over their gills. In some fish, capillary blood flows in the opposite direction to the water, causing countercurrent exchange. The gills push the oxygen-poor water out through openings in the sides of the pharynx. Some newly hatched fry have undeveloped gills when they first hatch. These fry are able to use a process called cutaneous respiration where they absorb enough oxygen through their skin to meet their needs until their gills develop.
Fish from multiple groups can live out of the water for extended time periods:
Most fishes will be satisfied with oxygen levels of 5 or 6 mg/l of free oxygen in their environment but if this level drops the fish will be seen gasping at the surface in an attempt to find the most oxygen.
Each filament contains a capillary network that provides a large surface area for exchanging oxygen and carbon dioxide. Fish exchange gases by pulling oxygen-rich water through their mouths and pumping it over their gills. In some fish, capillary blood flows in the opposite direction to the water, causing countercurrent exchange. The gills push the oxygen-poor water out through openings in the sides of the pharynx. Some newly hatched fry have undeveloped gills when they first hatch. These fry are able to use a process called cutaneous respiration where they absorb enough oxygen through their skin to meet their needs until their gills develop.
Fish from multiple groups can live out of the water for extended time periods:
- Amphibious fish such as the mudskipper can live and move about on land for up to several days, or live in stagnant or otherwise oxygen depleted water. Many such fish can breathe air via a variety of mechanisms.
- The skin of anguillid eels and may absorb oxygen directly.
- The buccal cavity of the electric eel may breathe air.
- Catfish of the families Loricariidae, Callichthyidae, and Scoloplacidae absorb air through their digestive tracts.
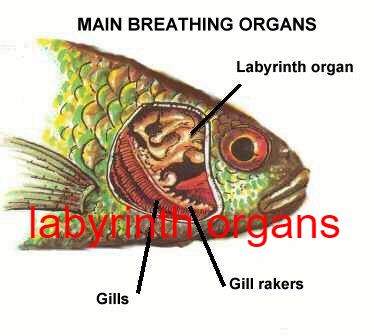
- A number of fish have evolved so-called accessory breathing organs that extract oxygen from the air (a labyrinth organ) as Labyrinth fish have a labyrinth organ above the gills that performs this function.

|
|

Air breathing fish can be divided into:
- Obligate air breathers
- Facultative air breathers

Facultative air breathers, such as the catfish Hypostomus plecostomus, only breathe air if they need to and can otherwise rely on their gills for oxygen.
Most air breathing fish are facultative air breathers that avoid the energetic cost of rising to the surface and the fitness cost of exposure to surface predators.
Bony fishIn bony fish, the gills lie in a branchial chamber covered by a bony operculum. The great majority of bony fish species have five pairs of gills. The operculum can be important in adjusting the pressure of water inside of the pharynx to allow proper ventilation of the gills. Valves inside the mouth keep the water from escaping.
Marine teleosts also use gills to excrete electrolytes.
- Saltwater fish lose large quantities of water osmotically through their gills. To regain the water, they drink large amounts of seawater and excrete the salt.
- Freshwater is more dilute than the internal fluids of fish, however, so freshwater fish gain water osmotically through their gills.
Cartilaginous fishLike other fish, sharks extract oxygen from seawater as it passes over their gills. Unlike other fish, shark gill slits are not covered, but lie in a row behind the head. A modified slit called a spiracle lies just behind the eye, which assists the shark with taking in water during respiration and plays a major role in bottom–dwelling sharks.

Pelagic Shark
Spiracles are reduced or missing in active pelagic sharks. While the shark is moving, water passes through the mouth and over the gills in a process known as "ram ventilation". A small number of species have lost the ability to pump water through their gills and must swim without rest. These species are obligate ram ventilators and would presumably asphyxiate if unable to move.
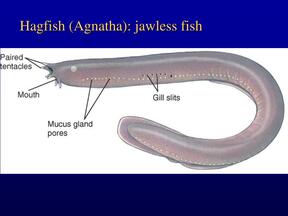
Lampreys and hagfishLampreys and hagfish do not have gill slits as such. Instead, the gills are contained in spherical pouches, with a circular opening to the outside. Like the gill slits of higher fish, each pouch contains two gills. In some cases, the openings may be fused together, effectively forming an operculum.
Lampreys have seven pairs of pouches, while hagfishes may have six to fourteen, depending on the species.
In the hagfish, the pouches connect with the pharynx internally. In adult lampreys, a separate respiratory tube develops beneath the pharynx proper, separating food and water from respiration by closing a valve at its anterior end.
Spiracles are reduced or missing in active pelagic sharks. While the shark is moving, water passes through the mouth and over the gills in a process known as "ram ventilation". A small number of species have lost the ability to pump water through their gills and must swim without rest. These species are obligate ram ventilators and would presumably asphyxiate if unable to move.
Lampreys and hagfishLampreys and hagfish do not have gill slits as such. Instead, the gills are contained in spherical pouches, with a circular opening to the outside. Like the gill slits of higher fish, each pouch contains two gills. In some cases, the openings may be fused together, effectively forming an operculum.
Lampreys have seven pairs of pouches, while hagfishes may have six to fourteen, depending on the species.
In the hagfish, the pouches connect with the pharynx internally. In adult lampreys, a separate respiratory tube develops beneath the pharynx proper, separating food and water from respiration by closing a valve at its anterior end.

The Circulation
Circulatory system and blood circulation is important for the survival in humans or animals.
For proper strategies of blood circulation management we need information on nutrients and waste products delivery in any animal which depends on circulatory system or diffusion from unicellular animal to multicellular animals respectively.
Function
The circulatory system performs many different functions:
- It is responsible for carrying oxygen to cells of the body.
- It also delivers nutrients to produce heat and energy.
- It acts as a delivery system for hormones to specific target organs.
- It performs by collecting metabolic waste and delivering it to excretory organs.
- It plays a protective role by combating infections and helping to establish an immune system defense.
- No circulatory Sys. animals.
- Open circulatory sys. animals.
- Closed circulat. sys. animals
Animals without circulatory system
These animals like the flatworms exhibit no circulatory system, but they are able to obtain nutrients, water, and oxygen by diffusion.
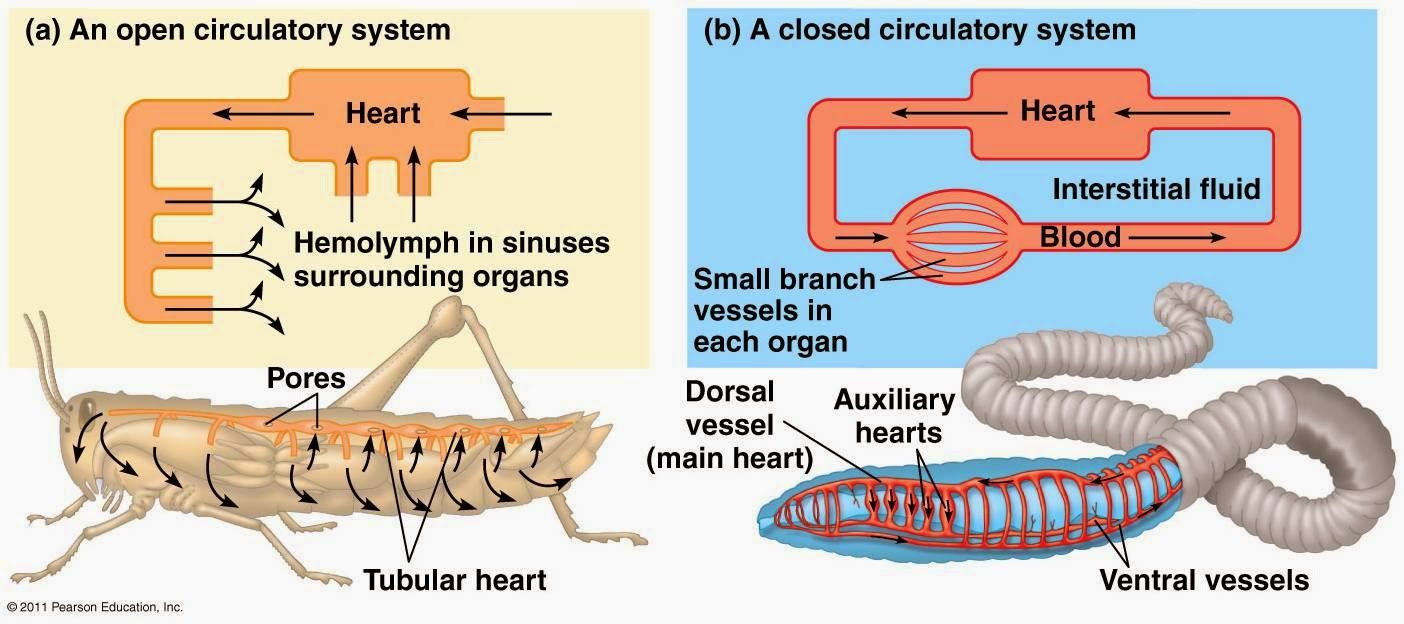
Animals with open circulatory system
· The open circulatory system can be found in arthropods and most mollusks and contains no capillaries or veins.
· In it a heart pumps (hemolymph) through arteries and into spaces around the organs.
· This allows tissues to exchange materials with the hemolymph, then drawn back into the heart as it relaxes.
Animals with closed circulatory system
·In the closed circulatory system, the blood remains inside the blood vessels and does not come out.
The blood flows from arteries to veins through small blood vessels called capillaries.
·
- The systems of fish, amphibians, reptiles, and birds show various stages of the evolution of the circulatory system.
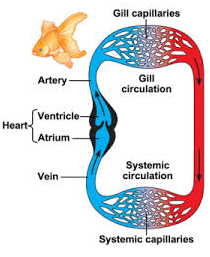
- In fish, the system has only one circuit, with the blood being pumped through the capillaries of the gills and on to the capillaries of the body tissues.
- This is known as single cycle circulation.
Blood Circulation In bony Fishes
- There are two kinds of capillary beds, those in the gills and those in the rest of the body.
- The heart, a folded continuous muscular tube with three or four sac-like enlargements, undergoes rhythmic contractions, and receives venous blood in a sinus venosus.
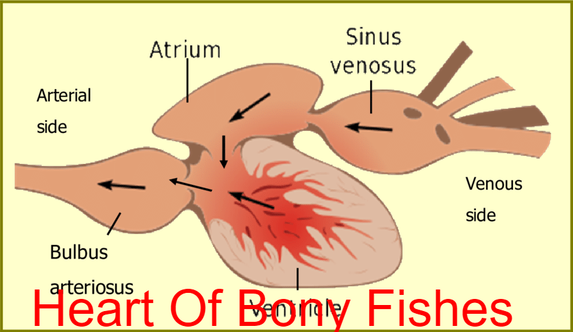
From the body To gill
- Auricle-------ventricle-------conus arteriosus-------ventral aorta------gills
- Afferent arteries of gill arches-----gill capillaries--------to body
Blood circulation in cartilaginous fishes
- In cartilaginous fishes a single circuit heart with 4four chambers; sinus venosus, atrium ventricle and conus arteriosus.
- The atrium is a thin-walled muscular sac; an A-V valve regulates flow between atrium & ventricle. The ventricle has thick, muscular walls. The conus arteriosus leads into the ventral aorta.
- A series of conal valves in the conus arteriosus prevent the backflow of blood.
- Ventral aorta extends forward below pharynx and connects developing aortic arches.
- Blood entering an aortic arch from ventral aorta must pass through gill capillaries before proceeding to dorsal aorta.
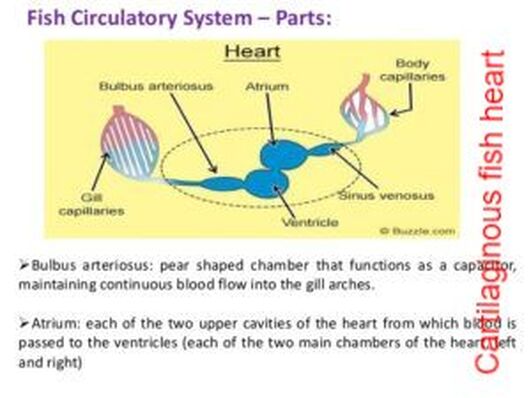
Differences between bony and cartilaginous fishes
Cartilaginous fishes (chondrichthyes):
Bony fishes (Osteichthyes):
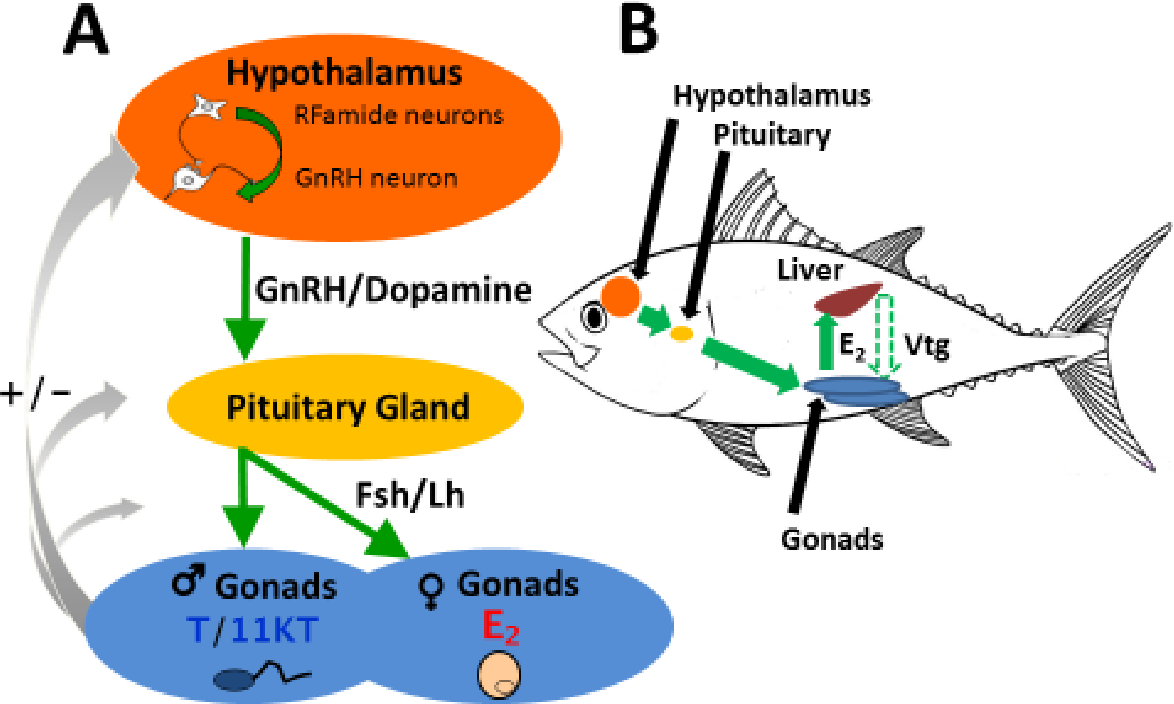
Fish Endocrinology
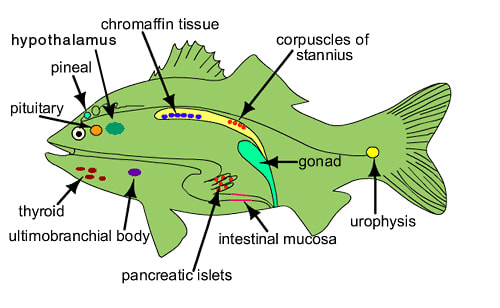
Fish endocrinology is limited to the work on its influence on chromatophores, action of sex cells, function of pituitary and thyroid and control on migration (Fig.). Endocrine glands are found in cyclostomes, elasmobranchs (sharks) and osteichthyes (bony fishes).
Cartilaginous fishes (chondrichthyes):
- Cartilaginous enoskeleton.
- Gills are exposed to the outside.
- Mouth on the ventral side of the head.
- Swim bladder absent.
- Placoid scales.
- Fertilization is internal.
- Examples; sharks and rays.
Bony fishes (Osteichthyes):
- Bony endoskeleton.
- Gills covered by operculum.
- Mouth at terminal end of the head.
- Swim bladder present.
- Cycloid and ctenoid scales.
- Fertilization is external.
- Examples; salmon, Flying fish and Sea horse.
Fish Endocrinology
Fish endocrinology is limited to the work on its influence on chromatophores, action of sex cells, function of pituitary and thyroid and control on migration (Fig.). Endocrine glands are found in cyclostomes, elasmobranchs (sharks) and osteichthyes (bony fishes).
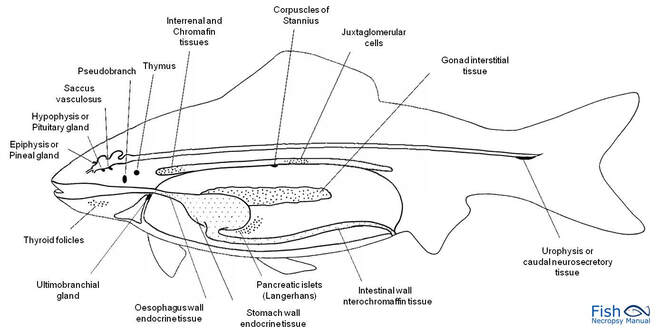
Corpuscle of Stannius
The corpuscles of of Stannius are found only in the kidney of bony fishes. They are responsible for maintaining calcium balance. Its hormone was identified as stanniocalcin. They were discovered and described by a German anatomist Hermann Friedrich Stannius (1839).
Stannius considered them as functionally similar to adrenal glands in mammals.
Chromaffin tissue
Chromaffin cells are generally dispersed in the head kidney near the post cardinal vein. Functional unit of adrenal medulla is chromaffin cells. Chromaffin tissue consists of two types of cells containing secretory granules, adrenaline and noreadrenaline cells. Named so because the cells are stained by solution containing chromium.
Chromaffin cells are generally dispersed in the head kidney near the post cardinal vein. Functional unit of adrenal medulla is chromaffin cells. Chromaffin tissue consists of two types of cells containing secretory granules, adrenaline and noreadrenaline cells. Named so because the cells are stained by solution containing chromium.
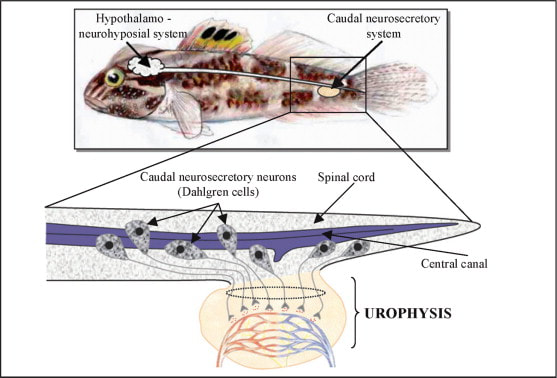
Urophysis
Urophysis is a small oval body, present in the terminal part of spinal cord . It is an organ, which releases materials produced in the neurosecretory cells situated in the spinal cord.
In some study, both neuropeptides isotocin (IT) and arginine vasotocin (AVT) have been identified and measured in urophysis, the neurohaemal organ of the caudal neurosecretory system of teleost fish.
Neuropeptide plays a different role in a circadian time keeping system and endocrine calendar in fish.
Urophysis is a small oval body, present in the terminal part of spinal cord . It is an organ, which releases materials produced in the neurosecretory cells situated in the spinal cord.
In some study, both neuropeptides isotocin (IT) and arginine vasotocin (AVT) have been identified and measured in urophysis, the neurohaemal organ of the caudal neurosecretory system of teleost fish.
Neuropeptide plays a different role in a circadian time keeping system and endocrine calendar in fish.
Ultimobranchial gland
Ultimobranchial gland is an endocrine gland producing calcitonin which helps regulate the calcium level. In bony fishes, the gland is located between the ventral wall of the oesophagus and the sinus venosus.
Hormones chemically can be divided into:
Ultimobranchial gland is an endocrine gland producing calcitonin which helps regulate the calcium level. In bony fishes, the gland is located between the ventral wall of the oesophagus and the sinus venosus.
Hormones chemically can be divided into:
- Steroid hormones (testosterone and estradiol)
- Protein hormones e.g. insulin .
- Amino acid hormones e.g thyroid hormones and epinephrine, norepinephrine (catecholamines).

Location of pituitary glands
This small pituitary gland can be found in the upper part of the fish head, on the ventral side of the brain.
The pituitary gland in fishes, as in all other vertebrates, is composed of the adeno-hypophysis and the neurohypophysis originating from the diencephalon (hypothalamus).
The adenohypophysis, which regulates gonadal functions in fishes, is the site of synthesis, storage and release into circulation of several peptide and protein hormones. The adenohypophysis is divided into the rostral (pro-adenohypophysis) and the proximal (meso-adenohypophysis) pars distalis, and the pars intermedia (meta-adenohypophysis).
The cells of pituitary secrete hormones and hormones are stored in granules present in the cytoplasm.
Recently on the basis of immunocytochemistry, the cells are classified according to hormones released by the pro-adenohypophysis.
For example, the cells which take basophilic stain produce adrenocorticotropic hormones, they are called ACTH cells but if secrete thyroid stimulating hormone, these cells are called thyrotrophs and if they secrete FSH hormones they are called gonadotrops.
Hormones of Pituitary Glands:
The different hormones secreting cell are not localized in specific region but are spread over in part of the adenohypophysis.
Section of pituitary gland showing various hormone secreting cells in the adenohypophysis. ACTH, adenocorticotropic cells; GTH, gonadotropic cells; MsA, mesoadenohypophysis; MtA, metaadenohypophysis; MSH melanotropic cells; Nh, neurohypophysis; PAS, periodic acid Schiff-positive cells in pars intermediate; P prolactin producing cell; STH, somatotropic cell; TSH ,thrytropic cell.
All hormones secreted by the pituitary are necessarily proteins or polypeptides. There exists a slight difference in the pituitary hormones of the different groups of fishes. The pituitary hormones of fishes are of two types (I) one which regulates the function of other endocrine glands. Such hormones are called tropins or tropic hormones.
These are:
Second which directly regulates the specific enzymatic reactions in the various body cells or tissues. These hormones are melanin hormones (MH) and melanophore stimulating hormone (MSH), etc. Thyrotropin hormone is secreted from pro-adenohypophysis (rostral pars distalis) and stimulate activity of thyroid hormones.
The TSH is secreted under the influence of (TRH), thyroid- releasing hormones from diencephalon in fishes. It is proved that TRH influences the TSH cell activity and thyroid activity in fish.
The gonadotropin (GTH) releasing hormone (GnRH) of teleost is similar to luteinizing hormone releasing hormone (LH-RH) is localized in ventral lateral nucleus preopticus periventricularis (NPP) and posterior lateral nucleus lateral tuberis (NLT) as well as other areas.
In several fishes GTH secretion is associated with ovulation. In Carassius auratus, GTH level becomes higher on the day of ovulation. However, in sockeye salmon, Oncorhynchus nerka, high level of GTH found during spawning.
In fishes there is only one functional gonadotropin is found, which is often regarded as piscian pituitary gonadotropin (PPG). This single gonadotropin has similar properties of two hormones, LH and FSH of mammals.
Adrenocorticotropic Hormone (ACTH):
It is secreted by ACTH cells located between the rostral pars distalis and the neurohypophysis. Secretion of ACTH from pituitary is stimulated by the hypothalamus through corticotrophin releasing factor (CRF).
Prolactin:
It is a similar hormone that influences lactation in mammals and is released from pro-adenohypophysis. In some fishes like mummichog (Fundulus heteroclitus), prolactin along with the intermedin enhances the laying down of melanin in the melanophores of the skin.
Growth Hormone (GH):
Mesoadenohypophysis secretes a growth hormone which accelerates increase in the body length of fishes. Very little is known concerning its control, mode of action on cell division and protein synthesis in teleosts.
Melanocyte Stimulating Hormone (MSH) or Intermedin:
MSH is secreted from the meta-adenohypophysis and acts antagonistically to melanin hormone (MAH). MSH expands the pigment in the chromatophores, thus takes part in adjustment of background. It also stimulates the melanin synthesis.
Oxytocoin and Vasopressin Hormones:
In fishes the neurohypophysis secretes two hormones, i.e., oxytocin and vasopressin, which are stored in hypothalmic neurosecretory cells. These endocrine substances have well known effect on mammalian metabolism.
The fish pituitary hormones are capable to produce such effects in higher vertebrates but presumably the target organs are specific site of their action in fishes and probably is different from those of higher vertebrates. In fishes they control osmoregulation by maintaining water and salt balance.
Thyroid Gland of Fishes
In many teleosts the thyroid gland is situated in the pharyngeal region in between the dorsal basibranchial cartilages and ventral sternohyoid muscle. The thyroid surrounds anterior and middle parts of first, second and sometimes third afferent branchial arteries of ventral aorta, as found in Ophiocephalus species.
In Heteropneustes it occupies almost the entire length of the ventral aorta and afferent arteries.
This small pituitary gland can be found in the upper part of the fish head, on the ventral side of the brain.
The pituitary gland in fishes, as in all other vertebrates, is composed of the adeno-hypophysis and the neurohypophysis originating from the diencephalon (hypothalamus).
The adenohypophysis, which regulates gonadal functions in fishes, is the site of synthesis, storage and release into circulation of several peptide and protein hormones. The adenohypophysis is divided into the rostral (pro-adenohypophysis) and the proximal (meso-adenohypophysis) pars distalis, and the pars intermedia (meta-adenohypophysis).
The cells of pituitary secrete hormones and hormones are stored in granules present in the cytoplasm.
Recently on the basis of immunocytochemistry, the cells are classified according to hormones released by the pro-adenohypophysis.
For example, the cells which take basophilic stain produce adrenocorticotropic hormones, they are called ACTH cells but if secrete thyroid stimulating hormone, these cells are called thyrotrophs and if they secrete FSH hormones they are called gonadotrops.
Hormones of Pituitary Glands:
The different hormones secreting cell are not localized in specific region but are spread over in part of the adenohypophysis.
Section of pituitary gland showing various hormone secreting cells in the adenohypophysis. ACTH, adenocorticotropic cells; GTH, gonadotropic cells; MsA, mesoadenohypophysis; MtA, metaadenohypophysis; MSH melanotropic cells; Nh, neurohypophysis; PAS, periodic acid Schiff-positive cells in pars intermediate; P prolactin producing cell; STH, somatotropic cell; TSH ,thrytropic cell.
All hormones secreted by the pituitary are necessarily proteins or polypeptides. There exists a slight difference in the pituitary hormones of the different groups of fishes. The pituitary hormones of fishes are of two types (I) one which regulates the function of other endocrine glands. Such hormones are called tropins or tropic hormones.
These are:
- Thyrotropin activates thyroid.
- Adrenocorticotropic hormones activate adrenal cortex.
- The gonadotropin FSH and LH (Leuteotropins, various steroid hormones).
- Growth hormones, somatotropin (actually they are not tropic).
Second which directly regulates the specific enzymatic reactions in the various body cells or tissues. These hormones are melanin hormones (MH) and melanophore stimulating hormone (MSH), etc. Thyrotropin hormone is secreted from pro-adenohypophysis (rostral pars distalis) and stimulate activity of thyroid hormones.
The TSH is secreted under the influence of (TRH), thyroid- releasing hormones from diencephalon in fishes. It is proved that TRH influences the TSH cell activity and thyroid activity in fish.
The gonadotropin (GTH) releasing hormone (GnRH) of teleost is similar to luteinizing hormone releasing hormone (LH-RH) is localized in ventral lateral nucleus preopticus periventricularis (NPP) and posterior lateral nucleus lateral tuberis (NLT) as well as other areas.
In several fishes GTH secretion is associated with ovulation. In Carassius auratus, GTH level becomes higher on the day of ovulation. However, in sockeye salmon, Oncorhynchus nerka, high level of GTH found during spawning.
In fishes there is only one functional gonadotropin is found, which is often regarded as piscian pituitary gonadotropin (PPG). This single gonadotropin has similar properties of two hormones, LH and FSH of mammals.
Adrenocorticotropic Hormone (ACTH):
It is secreted by ACTH cells located between the rostral pars distalis and the neurohypophysis. Secretion of ACTH from pituitary is stimulated by the hypothalamus through corticotrophin releasing factor (CRF).
Prolactin:
It is a similar hormone that influences lactation in mammals and is released from pro-adenohypophysis. In some fishes like mummichog (Fundulus heteroclitus), prolactin along with the intermedin enhances the laying down of melanin in the melanophores of the skin.
Growth Hormone (GH):
Mesoadenohypophysis secretes a growth hormone which accelerates increase in the body length of fishes. Very little is known concerning its control, mode of action on cell division and protein synthesis in teleosts.
Melanocyte Stimulating Hormone (MSH) or Intermedin:
MSH is secreted from the meta-adenohypophysis and acts antagonistically to melanin hormone (MAH). MSH expands the pigment in the chromatophores, thus takes part in adjustment of background. It also stimulates the melanin synthesis.
Oxytocoin and Vasopressin Hormones:
In fishes the neurohypophysis secretes two hormones, i.e., oxytocin and vasopressin, which are stored in hypothalmic neurosecretory cells. These endocrine substances have well known effect on mammalian metabolism.
The fish pituitary hormones are capable to produce such effects in higher vertebrates but presumably the target organs are specific site of their action in fishes and probably is different from those of higher vertebrates. In fishes they control osmoregulation by maintaining water and salt balance.
Thyroid Gland of Fishes
In many teleosts the thyroid gland is situated in the pharyngeal region in between the dorsal basibranchial cartilages and ventral sternohyoid muscle. The thyroid surrounds anterior and middle parts of first, second and sometimes third afferent branchial arteries of ventral aorta, as found in Ophiocephalus species.
In Heteropneustes it occupies almost the entire length of the ventral aorta and afferent arteries.
In Clarias batrachus the thyroid gland is concentrated around the ventral aorta, middle ends of two pairs of afferent arteries and the paired inferior jugular veins.
In lampreys, like higher fish, the follicles of thyroid glands are concentrated in the ventral pharynx and have an enough blood supply.
Hormones of Thyroid Gland:
Thyroid hormone is synthesized in the thyroid gland, for which inorganic iodine is extracted from the blood. These inorganic iodine combines with tyrosine. The thyroid hormones of fishes appear to be identical with those of mammals, including, mono- and di-iodo-tyrosine and thyroxin.
These hormones are kept stored in the thyroid follicles and are released into blood stream on metabolic demands. The release of the thyroid hormone from the follicle is controlled by the thyrotropic hormone (TSH) of pituitary which in turn is influenced by genetically determined maturation process along with certain factors like temperature, photoperiod and salinity. The thyroid glands in sharks and higher teleosts are diffused in nature.
In salmon the thyroid gland becomes hyperactive during their spawning. It has been considered that thyroid influences the growth and nitrogen metabolism in goldfish, as indicated by high ammonia excreted by them. Thus the action of thyroid is conjugated with other vital processes including maturation and also the diadromous migration of fishes.
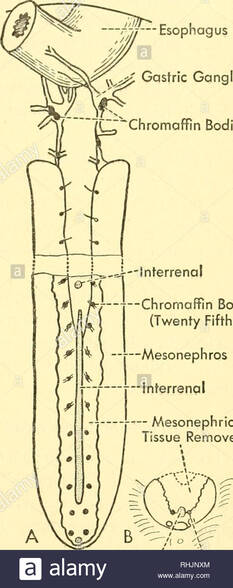
Adrenal Cortical Tissue ( Inter-Renal Tissue ):
In Lamprey (Cyclostomata) the endocrine inter-renal cells are present throughout the body cavity close to the post-cardinal vein. Among the rays they lie in more or less close association with posterior kidney tissue, including some species possessing inter-renal tissue concentrated near the left and in other near the right central border of that organ.
In sharks (Squaliformes) they are present between the kidneys. In teleosts the inter-renal cells are multilayered and situated along the post-cardinal veins as they enter the head kidney .
In lampreys, like higher fish, the follicles of thyroid glands are concentrated in the ventral pharynx and have an enough blood supply.
Hormones of Thyroid Gland:
Thyroid hormone is synthesized in the thyroid gland, for which inorganic iodine is extracted from the blood. These inorganic iodine combines with tyrosine. The thyroid hormones of fishes appear to be identical with those of mammals, including, mono- and di-iodo-tyrosine and thyroxin.
These hormones are kept stored in the thyroid follicles and are released into blood stream on metabolic demands. The release of the thyroid hormone from the follicle is controlled by the thyrotropic hormone (TSH) of pituitary which in turn is influenced by genetically determined maturation process along with certain factors like temperature, photoperiod and salinity. The thyroid glands in sharks and higher teleosts are diffused in nature.
In salmon the thyroid gland becomes hyperactive during their spawning. It has been considered that thyroid influences the growth and nitrogen metabolism in goldfish, as indicated by high ammonia excreted by them. Thus the action of thyroid is conjugated with other vital processes including maturation and also the diadromous migration of fishes.
Adrenal Cortical Tissue ( Inter-Renal Tissue ):
In Lamprey (Cyclostomata) the endocrine inter-renal cells are present throughout the body cavity close to the post-cardinal vein. Among the rays they lie in more or less close association with posterior kidney tissue, including some species possessing inter-renal tissue concentrated near the left and in other near the right central border of that organ.
In sharks (Squaliformes) they are present between the kidneys. In teleosts the inter-renal cells are multilayered and situated along the post-cardinal veins as they enter the head kidney .
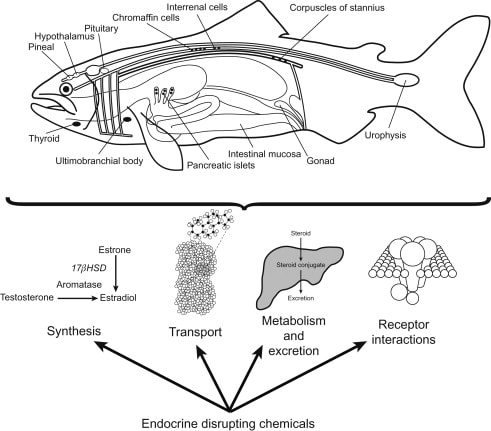
Adrenal Cortical Hormone:
Adrenal cortical tissue or inter-renal tissue secretes two hormones. These are (i) mineral corticoids concerned with fish osmoregulation.
(ii) glucocorticoids, which regulate the carbohydrate metabolism, particularly blood sugar level.
Salmo gairdneri treated with mineral corticoid excretes higher than normal amount of sodium ions through its gills but conserve more than normal amount of sodium in the kidneys and osmoregulation in the body.
Chromaffin Tissue ( Medullary Tissue ):
In lamprey (Cyclostomata) the chromaffin cells are present in the form of strands along the dorsal aorta as in the ventricle and the portal vein heart. In sharks and rays (Elasmobranchii) these tissues are found associated with the sympathetic chain of nerve ganglia while in bony fishes (Actinopterygii) the chromaffin cells have wide variation in their distribution.
The Ultimo-Branchial Gland:
Typically the gland is small and paired and is situated in the transverse septum between the abdominal cavity and sinus venosus just ventral to the oesophagus or near the thyroid gland.
The gland secretes the hormone calcitonin which regulates calcium metabolism.
Calcitonin is said to be related with the osmoregulation. Eel calcitonin causes decrease serum osmolarity, sodium and chloride in Japanese eels. The ultimo-branchial gland is under the control of pituitary gland.
The Sex Endocrine Glands:
The sex hormones are synthesized and secreted by specialized cells of the ovaries and testis. The release of sex hormones are under the control of mesoadenohypophysis of pituitary. In fishes these sex hormones are necessary for maturation of gametes and in addition secondary sex characteristics such as breeding tubercles, colouration and the maturation of gonopodia.
In elasmobranch (Raja) and in salmon the blood plasma contains male hormone testosterone with a correlation between plasma level and the reproductive cycle. Oryzias latipes (medaka) and sockey salmon comprise another gonadal steroid, i.e., 11-ketotestosteron, which is 10 fold more physiologically androgenic than testosterone.
Intestinal Mucosa:
The intestinal mucosa produces secretin and pancreozymin, which are controlled by nervous system and regulate pancreatic secretion. Secretin affects flow of enzyme carrying liquids from the pancreas, whereas pancreozymin accelerates flow of zymogens.
These hormones are usually synthesized in anterior part of the small intestine. In carnivorous fish these hormones are brought into the stomach, containing acidified homogenate of fish flesh or by injection of secretin into gastric vein which stimulates the secretion of pancreas.
Islets of Langerhans:
In some fishes like Labeo, Cirrhina, and Channa small islets are present which are separate from pancreas and are found near gall bladder, spleen, pyloric caeca or intestine. Such islets are often referred to as principal islets. But in some species like Clarias batrachus and Heteropneustes fossils the number of large and small islets are found to be embedded in the pancreatic tissues, similar to the higher vertebrates.
Two kinds of fish islets cells:
(i) The beta cells which secrete insulin and take aldehyde fuschin stain.
(ii) Another type of cells are alpha cells, which do not take aldehyde fuschin stain and have two types, A1 and A2 cells, which produce glucagon. The function of the third type of cells is not known. Insulin is secreted by beta cells and regulates the blood sugar level in fishes.
Pineal Organ:
It is situated near the pituitary. In-spite of being a photoreceptor organ the pineal organ shows endocrine nature of doubtful function. Removal of pineal from Lebistes species causes reduced growth rate, anomalies in the skeleton, pituitary, thyroid and corpuscles of Stannius. It has been reported that thyroid and pituitary glands influence the secretion of pineal.
---------------------------------------------------
The Digestion in Fish
Jaws allow fish to eat a wide variety of food, including plants and other organisms. Fish ingest food through the mouth and break it down in the esophagus. In the stomach, food is further digested and, in many fish, processed in finger-shaped pouches called pyloric caeca, which secrete digestive enzymes and absorb nutrients. Organs such as the liver and pancreas add enzymes and various chemicals as the food moves through the digestive tract. The intestine completes the process of digestion and nutrient absorption.
In most vertebrates, digestion is a four-stage process involving the main structures of the digestive tract, starting with ingestion, placing food into the mouth, and concluding with the excretion of undigested material through the anus. From the mouth, the food moves to the stomach, where as bolus it is broken down chemically. It then moves to the intestine, where the process of breaking the food down into simple molecules continues and the results are absorbed as nutrients into the circulatory and lymphatic system.
Although the precise shape and size of the stomach varies widely among different vertebrates, the relative positions of the oesophageal and duodenal openings remain relatively constant. As a result, the organ always curves somewhat to the left before curving back to meet the pyloric sphincter. However, lampreys, hagfishes, chimaeras, lungfishes, and some teleost fish have no stomach at all, with the oesophagus opening directly into the intestine. These animals all consume diets that either require little storage of food, or no pre-digestion with gastric juices, or both.
The small intestine is the part of the digestive tract following the stomach and followed by the large intestine, and is where much of the digestion and absorption of food takes place. In fish, the divisions of the small intestine are not clear, and the terms anterior or proximal intestine may be used instead of duodenum. The small intestine is found in all teleosts, although its form and length vary enormously between species. In teleosts, it is relatively short, typically around one and a half times the length of the fish's body. It commonly has a number of pyloric caeca, small pouch-like structures along its length that help to increase the overall surface area of the organ for digesting food. There is no ileocaecal valve in teleosts, but the boundary between the small intestine and the rectum being marked only by the end of the digestive epithelium.
There is no small intestine as such in non-teleost fish, such as sharks, sturgeons, and lungfish. Instead, the digestive part of the gut forms a spiral intestine, connecting the stomach to the rectum. In this type of gut, the intestine itself is relatively straight, but has a long fold running along the inner surface in a spiral fashion, sometimes for dozens of turns. This valve greatly increases both the surface area and the effective length of the intestine. The lining of the spiral intestine is similar to that of the small intestine in teleosts and non-mammalian tetrapods. In lampreys, the spiral valve is extremely small, possibly because their diet requires little digestion. Hagfish have no spiral valve at all, with digestion occurring for almost the entire length of the intestine, which is not subdivided into different regions.
The large intestine is the last part of the digestive system normally found in vertebrate animals. Its function is to absorb water from the remaining indigestible food matter, and then to pass useless waste material from the body. In fish, there is no true large intestine, but simply a short rectum connecting the end of the digestive part of the gut to the cloaca. In sharks, this includes a rectal gland that secretes salt to help the animal maintain osmotic balance with the seawater. The gland somewhat resembles a caecum in structure, but is not a homologous structure.
As with many aquatic animals, most fish release their nitrogenous wastes as ammonia. Some of the wastes diffuse through the gills. Blood wastes are filtered by the kidneys.
Saltwater fish tend to lose water because of osmosis. Their kidneys return water to the body. The reverse happens in freshwater fish: they tend to gain water osmotically. Their kidneys produce dilute urine for excretion. Some fish have specially adapted kidneys that vary in function, allowing them to move from freshwater to saltwater.
In sharks, digestion can take a long time. The food moves from the mouth to a J-shaped stomach, where it is stored and initial digestion occurs. Unwanted items may never get past the stomach, and instead the shark either vomits or turns its stomachs inside out and ejects unwanted items from its mouth. One of the biggest differences between the digestive systems of sharks and mammals is that sharks have much shorter intestines. This short length is achieved by the spiral valve with multiple turns within a single short section instead of a long tube-like intestine. The valve provides a long surface area, requiring food to circulate inside the short gut until fully digested, when remaining waste products pass into the cloaca.
Organs such as the liver and pancreas add enzymes and various chemicals as the food moves through the digestive tract. The intestine completes the process of digestion and nutrient absorption.
In most vertebrates, digestion is a four-stage process involving the main structures of the digestive tract, starting with ingestion, placing food into the mouth. From the mouth, the food moves to the stomach, where as bolus it is broken down chemically. It then moves to the intestine, where the process of breaking the food down into simple molecules continues and the results are absorbed as nutrients into the circulatory and lymphatic system.
Some Fishes have no stomach:
However, lampreys, hagfishes, chimaeras, lungfishes, and some teleost fish have no stomach at all, with the oesophagus opening directly into the intestine. These animals all consume diets that either require little storage of food, or no pre-digestion with gastric juices, or both.
The small intestine is the part of the digestive tract following the stomach and followed by the large intestine, and is where much of the digestion and absorption of food takes place.
In fish, the divisions of the small intestine are not clear, and the terms anterior or proximal intestine may be used instead of duodenum. The small intestine is found in all teleosts, although its form and length vary enormously between species. In teleosts, it is relatively short, typically around one and a half times the length of the fish's body.
The pyloric caeca secrete digestive enzymes and increase the surface area in the stomach for nutrient absorption.
It commonly has a number of pyloric caeca, small pouch-like structures along its length that help to increase the overall surface area of the organ for digesting food.
There is no ileocaecal valve in teleosts, with the boundary between the small intestine and the rectum being marked only by the end of the digestive epithelium.
In lampreys, the spiral valve is extremely small, possibly because their diet requires little digestion.
The large intestine is the last part of the digestive system normally found in vertebrate animals. Its function is to absorb water from the remaining indigestible food matter, and then to pass useless waste material from the body. In fish, there is no true large intestine, but simply a short rectum connecting the end of the digestive part of the gut to the cloaca. In sharks, this includes a rectal gland that secretes salt to help the animal maintain osmotic balance with the seawater. The gland somewhat resembles a caecum in structure, but is not a homologous structure.
As with many aquatic animals, most fish release their nitrogenous wastes as ammonia. Some of the wastes diffuse through the gills. Blood wastes are filtered by the kidneys.
Saltwater fish tend to lose water because of osmosis. Their kidneys return water to the body. The reverse happens in freshwater fish: they tend to gain water osmotically. Their kidneys produce dilute urine for excretion. Some fish have specially adapted kidneys that vary in function, allowing them to move from freshwater to saltwater.
In sharks, digestion can take a long time. The food moves from the mouth to a J-shaped stomach, where it is stored and initial digestion occurs. Unwanted items may never get past the stomach, and instead the shark either vomits or turns its stomachs inside out and ejects unwanted items from its mouth. One of the biggest differences between the digestive systems of sharks and mammals is that sharks have much shorter intestines. This short length is achieved by the spiral valve with multiple turns within a single short section instead of a long tube-like intestine. The valve provides a long surface area, requiring food to circulate inside the short gut until fully digested, when remaining waste products pass into the cloaca.
Organs such as the liver and pancreas add enzymes and various chemicals as the food moves through the digestive tract. The intestine completes the process of digestion and nutrient absorption.
In most vertebrates, digestion is a four-stage process involving the main structures of the digestive tract, starting with ingestion, placing food into the mouth. From the mouth, the food moves to the stomach, where as bolus it is broken down chemically. It then moves to the intestine, where the process of breaking the food down into simple molecules continues and the results are absorbed as nutrients into the circulatory and lymphatic system.
Some Fishes have no stomach:
However, lampreys, hagfishes, chimaeras, lungfishes, and some teleost fish have no stomach at all, with the oesophagus opening directly into the intestine. These animals all consume diets that either require little storage of food, or no pre-digestion with gastric juices, or both.
The small intestine is the part of the digestive tract following the stomach and followed by the large intestine, and is where much of the digestion and absorption of food takes place.
In fish, the divisions of the small intestine are not clear, and the terms anterior or proximal intestine may be used instead of duodenum. The small intestine is found in all teleosts, although its form and length vary enormously between species. In teleosts, it is relatively short, typically around one and a half times the length of the fish's body.
The pyloric caeca secrete digestive enzymes and increase the surface area in the stomach for nutrient absorption.
It commonly has a number of pyloric caeca, small pouch-like structures along its length that help to increase the overall surface area of the organ for digesting food.
There is no ileocaecal valve in teleosts, with the boundary between the small intestine and the rectum being marked only by the end of the digestive epithelium.
In lampreys, the spiral valve is extremely small, possibly because their diet requires little digestion.
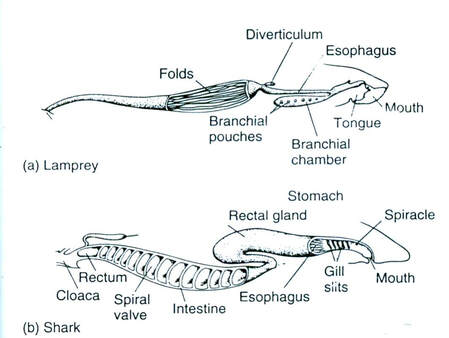
In Hagfish have no spiral valve at all, with digestion occurring for almost the entire length of the intestine, which is not subdivided into different regions.
The large intestine is the last part of the digestive system normally found in vertebrate animals. Its function is to absorb water from the remaining indigestible food matter, and then to pass useless waste material from the body.
In sharks, this includes a rectal gland that secretes salt to help the animal maintain osmotic balance with the seawater. The gland somewhat resembles a caecum in structure, but is not a homologous structure.
As with many aquatic animals, most fish release their nitrogenous wastes as ammonia. Some of the wastes diffuse through the gills. Blood wastes are filtered by the kidneys.
Saltwater fish tend to lose water because of osmosis. Their kidneys return water to the body.
The reverse happens in freshwater fish: they tend to gain water osmotically. Their kidneys produce dilute urine for excretion.
Some fish have specially adapted kidneys that vary in function, allowing them to move from freshwater to saltwater.
In sharks, digestion can take a long time. The food moves from the mouth to a J-shaped stomach, where it is stored and initial digestion occurs.
Unwanted materials may never get past the stomach, and instead the shark either vomits or turns its stomachs inside out and ejects unwanted materials from its mouth. One of the biggest differences between the digestive systems of sharks and mammals is that sharks have much shorter intestines.
The valve provides a long surface area, requiring food to circulate inside the short gut until fully digested, when remaining waste products pass into the cloaca.
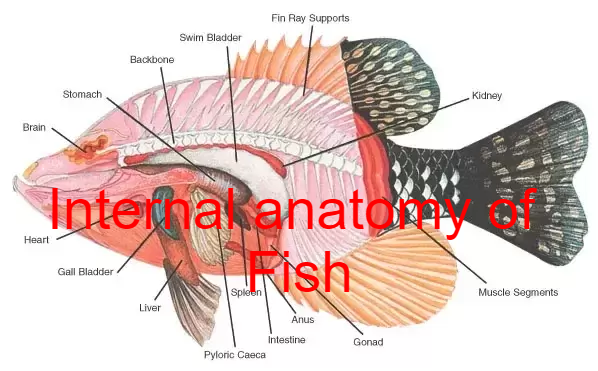
Liver
The liver is a large vital organ present in all fish. It has a wide range of functions, including detoxification, protein synthesis, and production of biochemicals necessary for digestion.
It is very susceptible to contamination by organic and inorganic compounds because they can accumulate over time and cause potentially life-threatening conditions. Because of the liver's capacity for detoxification and storage of harmful components, it is often used as an environmental biomarker.
Gall Bladder
The functions of bile have scarcely been studied in fish, but presumably resemble those in higher vertebrates. In mammals bile is composed mainly of bilirubin and biliverdin, which are breakdown products of haemoglobin, and is produced continuously. These salts act like detergents and serve to emulsify lipids, thus making lipids more accessible to enzymes because of the increased surface area, allowing some lipids to be absorbed undigested as micro-droplets.
That bile is produced continuously in fish is suggested by the presence of green mucus in the lumen of the atrophied gut of spawning salmon. There appear to be no studies in fish of gall bladder contraction or other mechanisms controlling the release of bile during digestion.
Spleen
The spleen is found in nearly all vertebrates. It is a non-vital organ, similar in structure to a large lymph node. It acts primarily as a blood filter, and plays important roles in regard to red blood cells and the immune system.
In cartilaginous and bony fish it consists primarily of red pulp and is normally a somewhat elongated organ as it actually lies inside the serosal lining of the intestine. The only vertebrates lacking a spleen are the lampreys and hagfishes. Even in lampreys and hagfishes, there is a diffuse layer of haematopoeitic tissue within the gut wall, which has a similar structure to red pulp, and is presumed to be homologous with the spleen of higher vertebrates.
N.B. Hematopoietic stem cells (HSCs) are the stem cells that give rise to blood cells through the process of haematopoiesis.
--------------------------------------------------------------
Fish Reproduction
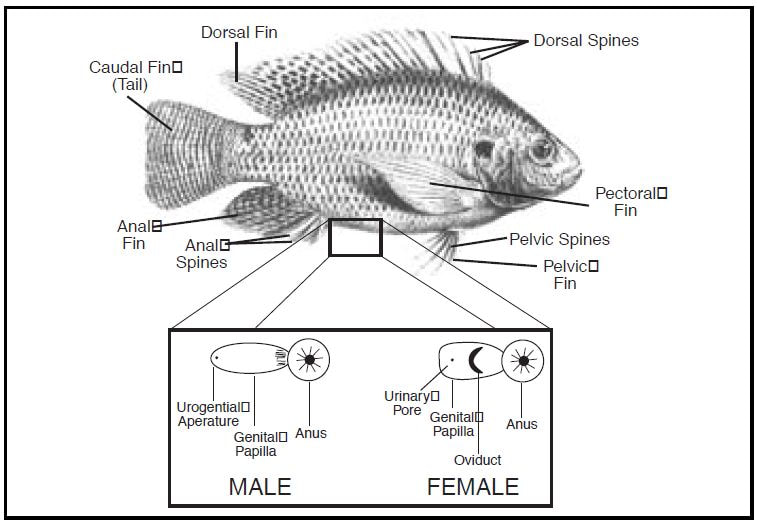
Fish reproductive organs include testes and ovaries. In most species, gonads are paired organs of similar size, which can be partially or totally fused. The genital papilla is a small, fleshy tube behind the anus in some fishes, from which the sperm or eggs are released.
TestesMost male fish have two testes of similar size. In the case of sharks, the testes on the right side is usually larger. The primitive jawless fish have only a single testis, located in the midline of the body.
Under a tough membranous shell, the tunica albuginea, the testis of some teleost fish, contains very fine coiled tubes called seminiferous tubules. The tubules are lined with a layer of cells (germ cells) that from puberty into old age, develop into sperm cells (also known as spermatozoa or male gametes).
The developing sperm travel through the seminiferous tubules to the efferent ducts, and then to the epididymis where newly created sperm cells mature (spermatogenesis). The sperm move into the vas deferens, and are eventually expelled through the urethra and out of the urethera through muscular contractions.
However, most fish do not possess seminiferous tubules. Instead, the sperm are produced in spherical structures called sperm ampullae. These are seasonal structures, releasing their contents during the breeding season, and then being reabsorbed by the body.
Ovaries
Many of the features found in ovaries are common to all vertebrates, including the presence of follicular cells and tunica albuginea. There may be hundreds or even millions of fertile eggs present in the ovary of a fish at any given time. Fresh eggs may be developing from the germinal epithelium throughout life. Corpora lutea are found only in mammals, and in some elasmobranch fish; in other species, the remnants of the follicle are quickly resorbed by the ovary.
The ovary of teleosts is often contains a hollow, lymph-filled space which opens into the oviduct, and into which the eggs are shed. Most normal female fish have two ovaries. In some elasmobranchs, only the right ovary develops fully. In the primitive jawless fish, and some teleosts, there is only one ovary, formed by the fusion of the paired organs in the embryo.
Fish ovaries may be of three types: primary gymnovarian, secondary gymnovarian or cystovarian. In the first type, the oocytes are released directly into the coelomic cavity and then enter the ostium, then through the oviduct and are eliminated. Secondary gymnovarian ovaries shed ova into the coelom from which they go directly into the oviduct. In the third type, the oocytes are conveyed to the exterior through the oviduct. Gymnovaries are the primitive condition found in lungfish, sturgeon, and bowfin. Cystovaries characterize most teleosts, where the ovary lumen has continuity with the oviduct. Secondary gymnovaries are found in salmonids and a few other teleosts.
The large intestine is the last part of the digestive system normally found in vertebrate animals. Its function is to absorb water from the remaining indigestible food matter, and then to pass useless waste material from the body.
In sharks, this includes a rectal gland that secretes salt to help the animal maintain osmotic balance with the seawater. The gland somewhat resembles a caecum in structure, but is not a homologous structure.
As with many aquatic animals, most fish release their nitrogenous wastes as ammonia. Some of the wastes diffuse through the gills. Blood wastes are filtered by the kidneys.
Saltwater fish tend to lose water because of osmosis. Their kidneys return water to the body.
The reverse happens in freshwater fish: they tend to gain water osmotically. Their kidneys produce dilute urine for excretion.
Some fish have specially adapted kidneys that vary in function, allowing them to move from freshwater to saltwater.
In sharks, digestion can take a long time. The food moves from the mouth to a J-shaped stomach, where it is stored and initial digestion occurs.
Unwanted materials may never get past the stomach, and instead the shark either vomits or turns its stomachs inside out and ejects unwanted materials from its mouth. One of the biggest differences between the digestive systems of sharks and mammals is that sharks have much shorter intestines.
The valve provides a long surface area, requiring food to circulate inside the short gut until fully digested, when remaining waste products pass into the cloaca.
Liver
The liver is a large vital organ present in all fish. It has a wide range of functions, including detoxification, protein synthesis, and production of biochemicals necessary for digestion.
It is very susceptible to contamination by organic and inorganic compounds because they can accumulate over time and cause potentially life-threatening conditions. Because of the liver's capacity for detoxification and storage of harmful components, it is often used as an environmental biomarker.
Gall Bladder
The functions of bile have scarcely been studied in fish, but presumably resemble those in higher vertebrates. In mammals bile is composed mainly of bilirubin and biliverdin, which are breakdown products of haemoglobin, and is produced continuously. These salts act like detergents and serve to emulsify lipids, thus making lipids more accessible to enzymes because of the increased surface area, allowing some lipids to be absorbed undigested as micro-droplets.
That bile is produced continuously in fish is suggested by the presence of green mucus in the lumen of the atrophied gut of spawning salmon. There appear to be no studies in fish of gall bladder contraction or other mechanisms controlling the release of bile during digestion.
Spleen
The spleen is found in nearly all vertebrates. It is a non-vital organ, similar in structure to a large lymph node. It acts primarily as a blood filter, and plays important roles in regard to red blood cells and the immune system.
In cartilaginous and bony fish it consists primarily of red pulp and is normally a somewhat elongated organ as it actually lies inside the serosal lining of the intestine. The only vertebrates lacking a spleen are the lampreys and hagfishes. Even in lampreys and hagfishes, there is a diffuse layer of haematopoeitic tissue within the gut wall, which has a similar structure to red pulp, and is presumed to be homologous with the spleen of higher vertebrates.
N.B. Hematopoietic stem cells (HSCs) are the stem cells that give rise to blood cells through the process of haematopoiesis.
--------------------------------------------------------------
Fish Reproduction
Fish reproductive organs include testes and ovaries. In most species, gonads are paired organs of similar size, which can be partially or totally fused. The genital papilla is a small, fleshy tube behind the anus in some fishes, from which the sperm or eggs are released.
TestesMost male fish have two testes of similar size. In the case of sharks, the testes on the right side is usually larger. The primitive jawless fish have only a single testis, located in the midline of the body.
Under a tough membranous shell, the tunica albuginea, the testis of some teleost fish, contains very fine coiled tubes called seminiferous tubules. The tubules are lined with a layer of cells (germ cells) that from puberty into old age, develop into sperm cells (also known as spermatozoa or male gametes).
The developing sperm travel through the seminiferous tubules to the efferent ducts, and then to the epididymis where newly created sperm cells mature (spermatogenesis). The sperm move into the vas deferens, and are eventually expelled through the urethra and out of the urethera through muscular contractions.
However, most fish do not possess seminiferous tubules. Instead, the sperm are produced in spherical structures called sperm ampullae. These are seasonal structures, releasing their contents during the breeding season, and then being reabsorbed by the body.
Ovaries
Many of the features found in ovaries are common to all vertebrates, including the presence of follicular cells and tunica albuginea. There may be hundreds or even millions of fertile eggs present in the ovary of a fish at any given time. Fresh eggs may be developing from the germinal epithelium throughout life. Corpora lutea are found only in mammals, and in some elasmobranch fish; in other species, the remnants of the follicle are quickly resorbed by the ovary.
The ovary of teleosts is often contains a hollow, lymph-filled space which opens into the oviduct, and into which the eggs are shed. Most normal female fish have two ovaries. In some elasmobranchs, only the right ovary develops fully. In the primitive jawless fish, and some teleosts, there is only one ovary, formed by the fusion of the paired organs in the embryo.
Fish ovaries may be of three types: primary gymnovarian, secondary gymnovarian or cystovarian. In the first type, the oocytes are released directly into the coelomic cavity and then enter the ostium, then through the oviduct and are eliminated. Secondary gymnovarian ovaries shed ova into the coelom from which they go directly into the oviduct. In the third type, the oocytes are conveyed to the exterior through the oviduct. Gymnovaries are the primitive condition found in lungfish, sturgeon, and bowfin. Cystovaries characterize most teleosts, where the ovary lumen has continuity with the oviduct. Secondary gymnovaries are found in salmonids and a few other teleosts.
Eggs
The eggs of fish are jellylike. Cartilagenous fish (sharks, skates, rays, chimaeras) eggs are fertilized internally and exhibit a wide variety of both internal and external embryonic development. Most fish species spawn eggs that are fertilized externally, typically with the male inseminating the eggs after the female lays them. These eggs do not have a shell and would dry out in the air.
The eggs of fish are jellylike. Cartilagenous fish (sharks, skates, rays, chimaeras) eggs are fertilized internally and exhibit a wide variety of both internal and external embryonic development. Most fish species spawn eggs that are fertilized externally, typically with the male inseminating the eggs after the female lays them. These eggs do not have a shell and would dry out in the air.



Intromittent organsMale cartilaginous fishes (sharks and rays), as well as the males of some live-bearing ray finned fishes, have fins that have been modified to function as intromittent organs, reproductive appendages which allow internal fertilization. In ray finned fish they are called gonopodiums or andropodiums, and in cartilaginous fish they are called claspers.
- A gonopodium is a modified anal fin (it looks rather like a thin rod) that allows male fish of the families Anablepidae and Poeciliidae to briefly hook into the vent of a female fish to deposit sperm.
- An andropodium is a short modified anal fin in males of certain species of live-bearing fish in the family Goodeidae. It is movable and used to impregnate females during mating.

Reproductive strategies
In fish, fertilisation of eggs can be either external or internal. In many species of fish, fins have been modified to allow Internal fertilisation. Similarly, development of the embryo can be external or internal, although some species show a change between the two at various stages of embryo development. Authors described reproductive strategies in terms of the development of the zygote and the interrelationship with the parents; there are five classifications:
OvuliparityOvuliparity means the female lays unfertilised eggs (ova), which must then be externally fertilised. Examples of ovuliparous fish include salmon, goldfish, cichlids, tuna and eels. In the majority of these species, fertilisation takes place outside the mother's body, with the male and female fish shedding their gametes into the surrounding water.
OviparityOviparity is where fertilisation occurs internally and so the female sheds zygotes into the water, often with important outer tissues added. Over 97% of all known fish are oviparous, In oviparous fish, internal fertilisation requires the male to use some sort of intromittent organ to deliver sperm into the genital opening of the female. Examples include the oviparous sharks, such as the horn shark, and oviparous rays, such as skates. In these cases, the male is equipped with a pair of claspers.
The newly hatched young of oviparous fish are called larvae. They are usually poorly formed, carry a large yolk sac and are very different in appearance from juvenile and adult specimens. The larval period in oviparous fish is relatively short (usually only several weeks), and larvae rapidly grow and change appearance and structure (a process termed metamorphosis) to become juveniles. During this transition larvae must switch from their yolk sac to feeding on zooplankton prey, a process which depends on typically inadequate zooplankton density, starving many larvae.
OvoviviparityIn ovoviviparous fish the eggs develop inside the mother's body after internal fertilisation but receive little or no nourishment directly from the mother, depending instead on a food reserve inside the egg, the yolk. Each embryo develops in its own egg. Familiar examples of ovoviviparous fish include guppies, angel sharks, and coelacanths.
ViviparityThere are two types of viviparity, differentiated by how the offspring gain their nutrients.
- Histotrophic (tissue eating) viviparity means embryos develop in the female’s oviducts but obtain nutrients by consuming other tissues, such as ova (oophagy) or zygotes. This has been observed primarily among sharks such as the shortfin mako and porbeagle, but is known for a few bony fish as well such as the halfbeak Nomorhamphus ebrardtii.
- Hemotrophic (blood eating) viviparity means embryos develop in the female’s (or male's) oviduct and nutrients are provided directly by the parent, typically via a structure similar to, or analogous to the placenta seen in mammals. Examples of hemotrophic fish include the surfperches, splitfins, lemon shark, seahorses and pipefish.
HermaphroditismHermaphroditism occurs when a given individual in a species possesses both male and female reproductive organs, or can alternate between possessing first one, and then the other. It can be contrasted with gonochorism, where each individual in a species is either male or female, and remains that way throughout their lives. Most fish are gonochorists, but hermaphroditism is known to occur in 14 families of teleost fishes.
Usually hermaphrodites are sequential, meaning they can switch sex, usually from female to male (protogyny). This can happen if a dominant male is removed from a group of females. The largest female can switch sex over a few days and replace the dominant male. This is found amongst coral reef fishes such as groupers, parrotfishes and wrasses. It is less common for a male to switch to a female (protandry).
Sexual parasitism
Sexual parasitism is a mode of sexual reproduction, unique to anglerfish, in which the males of a species are much smaller than the females, and rely on the females for food and protection from predators. The males give nothing back except the sperm which the females need in order to produce the next generation.
When a male finds a female, he bites into her skin, and releases an enzyme that digests the skin of his mouth and her body, fusing the pair down to the blood-vessel level. The male becomes dependent on the female host for survival by receiving nutrients via their now-shared circulatory system, and provides sperm to the female in return. After fusing, males increase in volume and become much larger relative to free-living males of the species. They live and remain reproductively functional as long as the female stays alive, and can take part in multiple spawnings. This extreme sexual dimorphism ensures that when the female is ready to spawn she has a mate immediately available. Multiple males can be incorporated into a single individual female with up to eight males in some species, though some types appear to have a one male per female.
ParthenogenesisParthenogenesis is a form of asexual reproduction in which growth and development of embryos occur without fertilization. In animals, parthenogenesis means development of an embryo from an unfertilized egg cell.
Parthenogenesis in sharks has been confirmed in the bonnethead and zebra shark. Other, usually sexual species, may occasionally reproduce parthenogenetically, and the hammerhead and blacktip sharks are recent additions to the known list of facultative parthenogenetic vertebrates.
-------------------------------------------------
Fish Metabolism
The oxygen consumption, or metabolic rate, of a fish at rest tells us about the amount of energy that fish requires to stay alive.
A fish’s metabolic requirements (or energy needs) will change at different points in its life. For example, a female Chinook salmon preparing to spawn needs to produce enough energy to survive and swim up river, while also using energy to produce her eggs. Fish metabolic rates are also affected by environmental conditions, such as temperature. Unlike in people, a fish’s body temperature will match the water surrounding it. Chemical reactions, like the ones in the process of metabolism, will speed up with warming temperatures, meaning a fish’s metabolic rate increases as the surrounding water temperature increases. In fact, increasing water temperature from 10°C to 20°C will cause the metabolic rate of a resting fish to double – that means the fish needs twice as much energy just to stay alive while resting! Environmental conditions can also impair metabolism. When water oxygen levels are low, fish can struggle to quickly uptake oxygen from the water. By limiting the rate of metabolism, low water oxygen levels can prevent a fish from performing some of the activities necessary for survival, like finding food, digesting meals, or escaping predators.
Because the ability of fish to perform the necessary activities for survival is key to determining where fish can live, environmental effects on metabolism influence fish habitat ranges. The researches developed a ‘metabolic index’, which essentially compares the water oxygen levels to the metabolic oxygen requirements of the fish. A low metabolic index indicates that a fish will have difficulty getting enough oxygen to meet its needs. The team predicted that most fish need a metabolic index of at least 2 to 5 in order to uptake enough oxygen to meet their energy requirements for survival.
Metabolic rate measurements can also be used in bioenergetics models, which compare the energy used through metabolism with the energy obtained through eating food to predict growth rates and food requirements of animals.
A fish’s metabolic rate can be very sensitive to its life stage and environment, and can influence fish behavior and survival. Thus, there are many important things we can learn from measuring the metabolic rate of fish under different conditions.
How quickly the animal consumes oxygen
When salmon and trout are exposed to very high temperatures, they often die. Thanks to the study of physiology, we now know that one cause of death in fish living at high temperatures can be a heart attack – fishes’ hearts try to speed up to accommodate the warm temperatures, but eventually can’t keep up. Physiology is the study of how an animal’s body functions and responds to its environment. In fishes, physiology studies often involve measuring factors such as swimming performance, heart rate, oxygen consumption, body chemistry and hormones, and survival under a variety of conditions. Knowledge of fish physiology is essential for managing fish populations, especially in artifitial waterways.
Aerobic scope measurements of fishes
Measurements of aerobic scope [the difference between minimum and maximum oxygen consumption rate ( and , respectively)] are increasing in prevalence as a tool to address questions relating to fish ecology and the effects of climate change.
The difference between a fish’s resting and swimming metabolic rates is called ‘aerobic scope’ because it is the scope or range of the fish’s ability to increase its metabolism. A steelhead trout’s aerobic scope is usually quite large, but becomes very small at high water temperatures. This is because at very warm temperatures, the heart reaches a point where it cannot speed up any more, even though the fish is just resting – the fish’s resting metabolic rate becomes almost as high as its swimming metabolic rate. Fish are in big trouble when their aerobic scope approaches zero because their hearts cannot pump enough to support swimming, eating, or escaping predators. In this study, we were able to identify the temperatures at which the fish’s aerobic scope became dangerously narrow. Knowing this relationship between water temperature and Central Valley steelhead and salmon aerobic scope can inform more efficient management of water resources to best protect the fish.
Such physiological studies can provide many valuable insights to inform fish conservation and management. These include how fishes respond to and tolerate a range of environmental conditions, and how fish bodies and internal processes change throughout different life stages (including smoltification and reproduction). Physiological studies can also reveal the extent to which individuals within and among populations vary in their physiological responses, and which stressors produce particular physiological consequences in fishes. Although population-level monitoring and tracking studies, such as surveying fish abundances, movements, and habitat use, are also essential tools for fish conservation, many important questions can only be addressed by studying physiology.
Physiological tools are particularly valuable for improving predictions of how fish populations will respond when faced with novel conditions. When creating management plans, scientists often look at population monitoring data to determine how fish populations responded to a set of environmental conditions in the past, and use this information to predict how a population will respond to contemporary or future conditions. However, without knowing the underlying mechanism that determined why populations responded in a certain way, we can only reliably use this approach to predict population responses under conditions that have been observed in the past. In contrast, physiological tools that measure the responses of individual fish to environmental changes can be combined with population monitoring to actually explain why population characteristics change, and can therefore be used to predict population responses to novel conditions in nature.
For example, a study on the metabolic responses of coral reef fishes to warm temperatures has provided essential knowledge for scientists’ predictions of how global warming will affect fish populations. Scientists have learned that marine species living near the equator are more susceptible to warming water temperatures due to global climate change than fish living at higher latitudes. This is because the warm equatorial water is already close to the highest temperatures that fish living there can tolerate.
Global climate change and rising human water demands make balancing the health of wildlife and the requirements of growing human populations increasingly challenging. Results of physiological studies are a powerful asset for developing management policies that allow both society and aquatic species to utilize river resources. However, when it comes to salmon and steelhead in the Central Valley, there are few studies related to the physiological effects of temperature, partly due to challenges of conducting on-site studies with wild fish, which limits our understanding of how these fish will respond to temperature changes. This lack of studies on local populations also means that temperature studies conducted in more northern and cooler climates are usually the basis for developing temperature tolerances and management criteria for southern populations, which may not be relevant.
Breathing Under Water
Oxygen is the key to life. All vertebrate animals require oxygen to live, and most of us cannot survive beyond a few short minutes without a generous oxygen supply. Water holds much less oxygen than air, yet some fish species can survive even in water with extremely low oxygen levels. The Crucian carp of Norway has evolved a particularly extraordinary ability to survive in small lakes and ponds that freeze over, becoming completely devoid of oxygen for months during the frigid northern European winter. What makes the Crucian carp truly remarkable is its special way to generate energy by converting the sugar glycogen into ethanol for survival when there is no oxygen in the water.
Scientists don’t think Crucian carp get tipsy from the ethanol they produce, but they can use ethanol to fill some of the same roles as oxygen in their metabolic pathways. However, this process is much less efficient than using oxygen, so the carp must reduce their energy demands – which they do by shutting down parts of the brain.
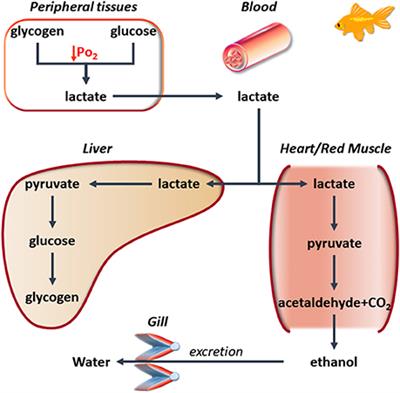
Some thick vegetative mats have devastating effects on oxygen levels in the river below. Dense populations of oxygen-consuming bacteria and fungi accumulate around hyacinth mats to feed on the constantly shedding plant material. These microorganisms consume the oxygen in the water, leaving little for fish.
Water oxygen concentrations of at least 6.0 mg/l are required for migrating salmon to survive, but levels were dropping as low as 2.0 mg/l. The good news is that a giant aeration system was constructed in 2007, which delivers up to 10,000 pounds of oxygen daily into the artifitial channel. This aeration system now maintains water oxygen levels above 5.2 mg/l through most of the year, and 6.2 mg/l when adult salmon are migrating past the channel.
Mobilization of glycogen stores was examined in the anoxic crucian carp (Carassius carassius Linnaeus). Winter-acclimatized fish were exposed to anoxia for 1, 3, or 6 weeks at 2 °C, and changes in the size of glycogen deposits were followed. After 1 week of anoxia, a major part of the glycogen stores was mobilized in liver (79.5 %) and heart (75.6 %), and large decreases occurred in gill (46.7 %) and muscle (45.1 %).
Brain was an exception in that its glycogen content remained unchanged. The amount of glycogen degraded during the first anoxic week was sufficient for the anaerobic ethanol production for more than 6 weeks of anoxia. After 3 and 6 weeks of anoxia, there was little further degradation of glycogen in other tissues except the brain where the stores were reduced by 30.1 and 49.9 % after 3 and 6 weeks of anoxia, respectively.
WATER CHEMISTRY
Dissolved Oxygen
Aquatic animals need dissolved oxygen (DO) to live. Fish, invertebrates, plants and aerobic bacteria all require oxygen for respiration. The amount of oxygen that can be dissolved in the water is reduced with increased temperature. The temperature effect is compounded by the fact that living organisms increase their activity in warm water, requiring more oxygen to support their metabolism. Fish and invertebrates that can move will leave areas with low DO.
DO levels < 3 ppm are stressful to most aquatic organisms.
DO levels < 2 ppm will not support fish.
DO = 5-6 ppm is required for growth and activity of most aquatic organisms.
Biological Oxygen Demand (BOD)
Oxygen is not only required for survival of most living organisms, but is needed to decompose organic (plant) material and human/animal wastes (sewage). BOD is determined by measuring the DO of a freshly collected sample and comparing it to the DO level in a sample that was collected at the same time but incubated in complete darkness, at 20°C, for 5 days. Unpolluted natural waters 5 mg/L BOD.
Nitrate-Nitrogen
Nitrogen is essential for plant growth, but the presence of excessive amounts in water supplies presents a major pollution problem. Nitrogen compounds may enter water from agricultural fertilizers, human sewage, industrial wastes, livestock wastes, and farm manure. Nitrate in drinking water must be 10 ppm.
pH
A pH of 6.5 to 8.2 is optimal for most organisms. Rapidly growing algae or submerged aquatic vegetation remove CO2 from the water during photosynthesis, significantly increasing pH levels. pH levels > 9.0 begin to be harmful to salmonids (trout) and perch. Rainwater naturally has a pH of 5.5; pH < 5.5 is harmful to freshwater shrimp, snails, and clams; metals normally trapped in sediments may be released into the acidified water.
Acidic < 6.5 pH Basic > 7.5 pH
Turbidity
Turbidity, or cloudiness in water, is caused by suspended materials that scatter light passing through the water. There are many possible sources of turbidity, including sediments from disturbed or eroded soil and high numbers of microscopic plankton due to excess nutrients and sunlight. Microscopic examination and streamwalk observations can help determine the sources of turbidity. In addition to blocking out the light needed by submerged aquatic vegetation and burying fish eggs and benthic (bottom dwelling) creatures, suspended sediment can carry nutrients and pesticides throughout the water system, damage gills, and interfere with the ability of fish to find food. Suspended particles near the water surface can absorb extra heat from sunlight, raising surface water temperatures.
Iron
Iron in water stains fixtures and may have an odor or taste. Values of 0-0.5 are acceptable. High values in streams may indicate contamination from landfills.
Phosphates
Phosphate test kits measure the form of phosphate applied as fertilizer to agricultural fields, grass lawns, or golf courses. Phosphates accelerate the growth of algae and aquatic plants. Total P > 0.03 ppm will increase plant growth and eutrophication.
Copper
Copper in water is from stamp sands or waste rock from copper mines. The national standard for aquatic life is .018 mg/l .
Alkalinity
Alkalinity is the amount of buffering material in the water. If a body of water has an abundance of buffering materials (high alkalinity), it is more stable and resistant to changes in pH. If a body of water has very little buffering material (low alkalinity), it is very susceptible to changes in pH. As increasing amounts of acid (acid rain) are added to ponds and lakes, their buffering capacity is consumed. If surrounding soils and rocks supply additional buffering materials, the alkalinity may eventually be restored. Even a temporary loss of buffering capacity can permit pH levels to drop to levels harmful to aquatic life.
Carbon Dioxide
Aquatic plants depend on CO2 in water for growth and respiration. CO2 increases when organic wastes reduce the oxygen available, making it difficult for fish to use the limited amount of oxygen present. Surface waters normally contain < 10 ppm of free CO2.
Water Temperatures
Temperature is one factor in determining which species may or may not be present. Temperature affects the feeding, reproduction, and metabolism of aquatic animals. A week or two of high temperature each year may make a stream unsuitable for sensitive aquatic organisms, even though temperatures are within tolerable levels throughout the rest of the year. Not only do different species have different temperature requirements, but the optimal temperature may change for different stages of life. Fish eggs and larvae usually have a narrower temperature require-ment than adult fish. Therefore, it is important to consider the life stage of species present when analyzing temperature data.
Water Color
The following list of possible colors and what they may be caused by:
Blue = transparent water with a low accumulation of dissolved materials and particulate matter,
indicates low productivity.
Yellow/Brown = dissolved organic materials, humic substances from soil, peat, or decaying plant material.
Red = can be produced by some algae.
Green = water rich in phytoplankton and other algae.
Mix of colors = may be caused by soil runoff.
--------------------------------------------------------------------------------------
Nervous System Of FishesCentral nervous system
Fish typically have quite small brains relative to body size compared with other vertebrates, typically one-fifteenth the brain mass of a similarly sized bird or mammal. However, some fish have relatively large brains, most notably mormyrids and sharks, which have brains about as massive relative to body weight as birds and marsupials.
Fish brains are divided into several regions. At the front are the olfactory lobes, a pair of structures that receive and process signals from the nostrilsvia the two olfactory nerves. The olfactory lobes are very large in fish that hunt primarily by smell, such as hagfish, sharks, and catfish. Behind the olfactory lobes is the two-lobed telencephalon, the structural equivalent to the cerebrum in higher vertebrates. In fish the telencephalon is concerned mostly with olfaction. Together these structures form the forebrain.
Connecting the forebrain to the midbrain is the diencephalon (in the diagram, this structure is below the optic lobes and consequently not visible). The diencephalon performs functions associated with hormones and homeostasis. The pineal body lies just above the diencephalon. This structure detects light, maintains circadian rhythms, and controls color changes. The midbrain (or mesencephalon) contains the two optic lobes. These are very large in species that hunt by sight, such as rainbow trout and cichlids.
Water oxygen concentrations of at least 6.0 mg/l are required for migrating salmon to survive, but levels were dropping as low as 2.0 mg/l. The good news is that a giant aeration system was constructed in 2007, which delivers up to 10,000 pounds of oxygen daily into the artifitial channel. This aeration system now maintains water oxygen levels above 5.2 mg/l through most of the year, and 6.2 mg/l when adult salmon are migrating past the channel.
Mobilization of glycogen stores was examined in the anoxic crucian carp (Carassius carassius Linnaeus). Winter-acclimatized fish were exposed to anoxia for 1, 3, or 6 weeks at 2 °C, and changes in the size of glycogen deposits were followed. After 1 week of anoxia, a major part of the glycogen stores was mobilized in liver (79.5 %) and heart (75.6 %), and large decreases occurred in gill (46.7 %) and muscle (45.1 %).
Brain was an exception in that its glycogen content remained unchanged. The amount of glycogen degraded during the first anoxic week was sufficient for the anaerobic ethanol production for more than 6 weeks of anoxia. After 3 and 6 weeks of anoxia, there was little further degradation of glycogen in other tissues except the brain where the stores were reduced by 30.1 and 49.9 % after 3 and 6 weeks of anoxia, respectively.
WATER CHEMISTRY
Dissolved Oxygen
Aquatic animals need dissolved oxygen (DO) to live. Fish, invertebrates, plants and aerobic bacteria all require oxygen for respiration. The amount of oxygen that can be dissolved in the water is reduced with increased temperature. The temperature effect is compounded by the fact that living organisms increase their activity in warm water, requiring more oxygen to support their metabolism. Fish and invertebrates that can move will leave areas with low DO.
DO levels < 3 ppm are stressful to most aquatic organisms.
DO levels < 2 ppm will not support fish.
DO = 5-6 ppm is required for growth and activity of most aquatic organisms.
Biological Oxygen Demand (BOD)
Oxygen is not only required for survival of most living organisms, but is needed to decompose organic (plant) material and human/animal wastes (sewage). BOD is determined by measuring the DO of a freshly collected sample and comparing it to the DO level in a sample that was collected at the same time but incubated in complete darkness, at 20°C, for 5 days. Unpolluted natural waters 5 mg/L BOD.
Nitrate-Nitrogen
Nitrogen is essential for plant growth, but the presence of excessive amounts in water supplies presents a major pollution problem. Nitrogen compounds may enter water from agricultural fertilizers, human sewage, industrial wastes, livestock wastes, and farm manure. Nitrate in drinking water must be 10 ppm.
pH
A pH of 6.5 to 8.2 is optimal for most organisms. Rapidly growing algae or submerged aquatic vegetation remove CO2 from the water during photosynthesis, significantly increasing pH levels. pH levels > 9.0 begin to be harmful to salmonids (trout) and perch. Rainwater naturally has a pH of 5.5; pH < 5.5 is harmful to freshwater shrimp, snails, and clams; metals normally trapped in sediments may be released into the acidified water.
Acidic < 6.5 pH Basic > 7.5 pH
Turbidity
Turbidity, or cloudiness in water, is caused by suspended materials that scatter light passing through the water. There are many possible sources of turbidity, including sediments from disturbed or eroded soil and high numbers of microscopic plankton due to excess nutrients and sunlight. Microscopic examination and streamwalk observations can help determine the sources of turbidity. In addition to blocking out the light needed by submerged aquatic vegetation and burying fish eggs and benthic (bottom dwelling) creatures, suspended sediment can carry nutrients and pesticides throughout the water system, damage gills, and interfere with the ability of fish to find food. Suspended particles near the water surface can absorb extra heat from sunlight, raising surface water temperatures.
Iron
Iron in water stains fixtures and may have an odor or taste. Values of 0-0.5 are acceptable. High values in streams may indicate contamination from landfills.
Phosphates
Phosphate test kits measure the form of phosphate applied as fertilizer to agricultural fields, grass lawns, or golf courses. Phosphates accelerate the growth of algae and aquatic plants. Total P > 0.03 ppm will increase plant growth and eutrophication.
Copper
Copper in water is from stamp sands or waste rock from copper mines. The national standard for aquatic life is .018 mg/l .
Alkalinity
Alkalinity is the amount of buffering material in the water. If a body of water has an abundance of buffering materials (high alkalinity), it is more stable and resistant to changes in pH. If a body of water has very little buffering material (low alkalinity), it is very susceptible to changes in pH. As increasing amounts of acid (acid rain) are added to ponds and lakes, their buffering capacity is consumed. If surrounding soils and rocks supply additional buffering materials, the alkalinity may eventually be restored. Even a temporary loss of buffering capacity can permit pH levels to drop to levels harmful to aquatic life.
Carbon Dioxide
Aquatic plants depend on CO2 in water for growth and respiration. CO2 increases when organic wastes reduce the oxygen available, making it difficult for fish to use the limited amount of oxygen present. Surface waters normally contain < 10 ppm of free CO2.
Water Temperatures
Temperature is one factor in determining which species may or may not be present. Temperature affects the feeding, reproduction, and metabolism of aquatic animals. A week or two of high temperature each year may make a stream unsuitable for sensitive aquatic organisms, even though temperatures are within tolerable levels throughout the rest of the year. Not only do different species have different temperature requirements, but the optimal temperature may change for different stages of life. Fish eggs and larvae usually have a narrower temperature require-ment than adult fish. Therefore, it is important to consider the life stage of species present when analyzing temperature data.
Water Color
The following list of possible colors and what they may be caused by:
Blue = transparent water with a low accumulation of dissolved materials and particulate matter,
indicates low productivity.
Yellow/Brown = dissolved organic materials, humic substances from soil, peat, or decaying plant material.
Red = can be produced by some algae.
Green = water rich in phytoplankton and other algae.
Mix of colors = may be caused by soil runoff.
--------------------------------------------------------------------------------------
Nervous System Of FishesCentral nervous system
Fish typically have quite small brains relative to body size compared with other vertebrates, typically one-fifteenth the brain mass of a similarly sized bird or mammal. However, some fish have relatively large brains, most notably mormyrids and sharks, which have brains about as massive relative to body weight as birds and marsupials.
Fish brains are divided into several regions. At the front are the olfactory lobes, a pair of structures that receive and process signals from the nostrilsvia the two olfactory nerves. The olfactory lobes are very large in fish that hunt primarily by smell, such as hagfish, sharks, and catfish. Behind the olfactory lobes is the two-lobed telencephalon, the structural equivalent to the cerebrum in higher vertebrates. In fish the telencephalon is concerned mostly with olfaction. Together these structures form the forebrain.
Connecting the forebrain to the midbrain is the diencephalon (in the diagram, this structure is below the optic lobes and consequently not visible). The diencephalon performs functions associated with hormones and homeostasis. The pineal body lies just above the diencephalon. This structure detects light, maintains circadian rhythms, and controls color changes. The midbrain (or mesencephalon) contains the two optic lobes. These are very large in species that hunt by sight, such as rainbow trout and cichlids.

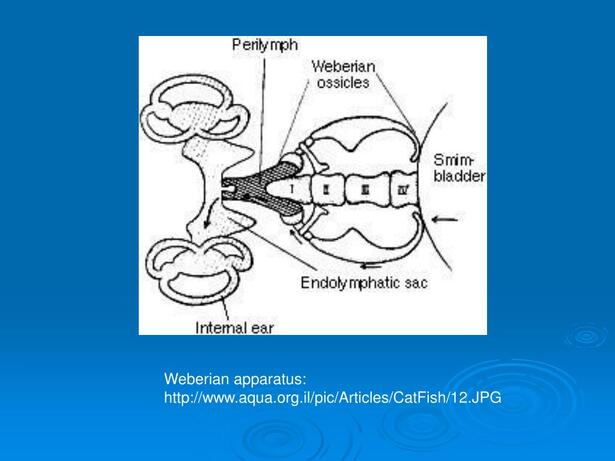
The hindbrain (or metencephalon) is particularly involved in swimming and balance. The cerebellum is a single-lobed structure that is typically the biggest part of the brain. Hagfish and lampreys have relatively small cerebellae, while the mormyrid cerebellum is massive and apparently involved in their electrical sense.
The brain stem (or myelencephalon) is the brain's posterior. As well as controlling some muscles and body organs, in bony fish at least, the brain stem governs respiration and osmoregulation.
Sensory Systems
Most fish possess highly developed sense organs. Nearly all daylight fish have color vision that is at least as good as a human's. Many fish also have chemoreceptors that are responsible for extraordinary senses of taste and smell. Although they have ears, many fish may not hear very well. Most fish have sensitive receptors that form the lateral line system, which detects gentle currents and vibrations, and senses the motion of nearby fish and prey. Sharks can sense frequencies in the range of 25 to 50 Hz through their lateral line.
Fish orient themselves using landmarks and may use mental maps based on multiple landmarks or symbols. Fish behavior in mazes متاهات reveals that they possess spatial memory and visual discrimination.
VisionVision is an important sensory system for most species of fish. Fish eyes are similar to those of terrestrial vertebrates like birds and mammals, but have a more spherical lens. Their retinas generally have both rod cells and cone cells (for scotopic and photopic vision), and most species have colour vision. Some fish can see ultraviolet and some can see polarized light. Amongst jawless fish, the lamprey has well-developed eyes, while the hagfish has only primitive eyespots. Fish vision shows adaptation to their visual environment, for example deep sea fishes have eyes suited to the dark environment.
HearingHearing is an important sensory system for most species of fish. Hearing threshold and the ability to localize sound sources are reduced underwater, in which the speed of sound is faster than in air. Underwater hearing is by bone conduction, and localization of sound appears to depend on differences in amplitude detected by bone conduction. Aquatic animals such as fish, however, have a more specialized hearing apparatus that is effective underwater.
Fish can sense sound through their lateral lines and their otoliths (ears). Some fishes, such as some species of carp and herring, hear through their swim bladders, which function rather like a hearing aid.
Hearing is well-developed in carp, which have the Weberian organ, three specialized vertebral processes that transfer vibrations in the swim bladder to the inner ear.
The brain stem (or myelencephalon) is the brain's posterior. As well as controlling some muscles and body organs, in bony fish at least, the brain stem governs respiration and osmoregulation.
Sensory Systems
Most fish possess highly developed sense organs. Nearly all daylight fish have color vision that is at least as good as a human's. Many fish also have chemoreceptors that are responsible for extraordinary senses of taste and smell. Although they have ears, many fish may not hear very well. Most fish have sensitive receptors that form the lateral line system, which detects gentle currents and vibrations, and senses the motion of nearby fish and prey. Sharks can sense frequencies in the range of 25 to 50 Hz through their lateral line.
Fish orient themselves using landmarks and may use mental maps based on multiple landmarks or symbols. Fish behavior in mazes متاهات reveals that they possess spatial memory and visual discrimination.
VisionVision is an important sensory system for most species of fish. Fish eyes are similar to those of terrestrial vertebrates like birds and mammals, but have a more spherical lens. Their retinas generally have both rod cells and cone cells (for scotopic and photopic vision), and most species have colour vision. Some fish can see ultraviolet and some can see polarized light. Amongst jawless fish, the lamprey has well-developed eyes, while the hagfish has only primitive eyespots. Fish vision shows adaptation to their visual environment, for example deep sea fishes have eyes suited to the dark environment.
HearingHearing is an important sensory system for most species of fish. Hearing threshold and the ability to localize sound sources are reduced underwater, in which the speed of sound is faster than in air. Underwater hearing is by bone conduction, and localization of sound appears to depend on differences in amplitude detected by bone conduction. Aquatic animals such as fish, however, have a more specialized hearing apparatus that is effective underwater.
Fish can sense sound through their lateral lines and their otoliths (ears). Some fishes, such as some species of carp and herring, hear through their swim bladders, which function rather like a hearing aid.
Hearing is well-developed in carp, which have the Weberian organ, three specialized vertebral processes that transfer vibrations in the swim bladder to the inner ear.
Although it is hard to test sharks' hearing, they may have a sharp sense of hearing and can possibly hear prey many miles away. A small opening on each side of their heads (not the spiracle) leads directly into the inner ear through a thin channel.
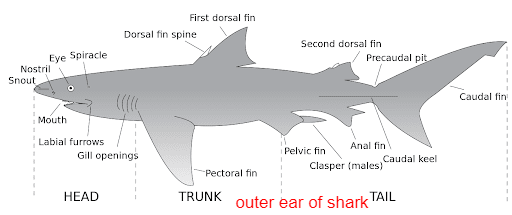
The lateral line shows a similar arrangement, and is open to the environment via a series of openings called lateral line pores. This is a reminder of the common origin of these two vibration- and sound-detecting organs that are grouped together as the acoustico-lateralis system. In bony fish and tetrapods the external opening into the inner ear has been lost.
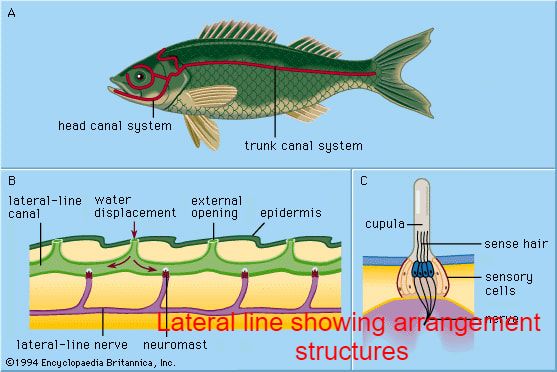
ChemoreceptionThe shape of the hammerhead shark's head may enhance olfaction by spacing the nostrils further apart.
Sharks have keen olfactory senses, located in the short duct (which is not fused, unlike bony fish) between the anterior and posterior nasal openings, with some species able to detect as little as one part per million of blood in seawater.
They are more attracted to the chemicals found in the intestines of many species, and as a result often linger near or in sewage outfalls. Some species, such as nurse sharks, have external barbels that greatly increase their ability to sense prey.
ElectroreceptionElectrocommunication
Electromagnetic field receptors (ampullae of Lorenzini) and motion detecting canals in the head of a shark.
Some fish, such as catfish and sharks, have organs that detect weak electric currents on the order of millivolt.
Other fish, like the South American electric fishes Gymnotiformes, can produce weak electric currents, which they use in navigation and social communication. In sharks, the ampullae of Lorenzini are electroreceptor organs. They number in the hundreds to thousands. Sharks use the ampullae of Lorenzini to detect the electromagnetic fields that all living things produce. This helps sharks (particularly the hammerhead shark) find prey. The shark has the greatest electrical sensitivity of any animal. Sharks find prey hidden in sand by detecting the electric fields they produce. Ocean currents moving in the magnetic field of the Earth also generate electric fields that sharks can use for orientation and possibly navigation.
Sharks have keen olfactory senses, located in the short duct (which is not fused, unlike bony fish) between the anterior and posterior nasal openings, with some species able to detect as little as one part per million of blood in seawater.
They are more attracted to the chemicals found in the intestines of many species, and as a result often linger near or in sewage outfalls. Some species, such as nurse sharks, have external barbels that greatly increase their ability to sense prey.
ElectroreceptionElectrocommunication
Electromagnetic field receptors (ampullae of Lorenzini) and motion detecting canals in the head of a shark.
Some fish, such as catfish and sharks, have organs that detect weak electric currents on the order of millivolt.
Other fish, like the South American electric fishes Gymnotiformes, can produce weak electric currents, which they use in navigation and social communication. In sharks, the ampullae of Lorenzini are electroreceptor organs. They number in the hundreds to thousands. Sharks use the ampullae of Lorenzini to detect the electromagnetic fields that all living things produce. This helps sharks (particularly the hammerhead shark) find prey. The shark has the greatest electrical sensitivity of any animal. Sharks find prey hidden in sand by detecting the electric fields they produce. Ocean currents moving in the magnetic field of the Earth also generate electric fields that sharks can use for orientation and possibly navigation.
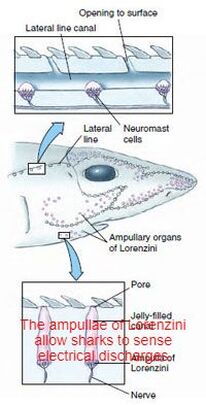
Electric fish are able to produce electric fields by modified muscles in their body.
Pain in fishExperiments done by William Tavolga provide evidence that fish have pain and fear responses. For instance, in Tavolga’s experiments, toadfish grunted when electrically shocked and over time they came to grunt at the mere sight of an electrode.
In 2003, Scottish scientists at the University of Edinburgh and the Roslin Institute concluded that rainbow trout exhibit behaviors often associated with pain as other animals. Bee venom and acetic acid injected into the lips resulted in fish rocking their bodies and rubbing their lips along the sides and floors of their tanks, which the researchers concluded were attempts to relieve pain, similar to what mammals would do. Neurons fired in a pattern resembling human neuronal patterns.
Professor James D. Rose of the University of Wyoming claimed the study was flawed since it did not provide proof that fish possess "conscious awareness, particularly a kind of awareness نوع من الوعي that is meaningfully like ours". Rose argues that since fish brains are so different from human brains, fish are probably not conscious in the manner humans are, so that reactions similar to human reactions to pain instead have other causes. Rose had published a study a year earlier arguing that fish cannot feel pain because their brains lack a neocortex.
Physiological Stress Response
In response to a stressor such as handling or crowding, a fish will undergo a series of biochemical and physiological changes in an attempt to compensate for the challenge imposed upon it and, thereby, cope with the stress. The stress response in fish has been broadly categorized into the primary, secondary, and tertiary responses.
Primary Response
This initial response represents the preception of an altered state and initiates a neuroendocrine/endocrine response that forms part of the generalized stress response in fish. This response includes the rapid release of stress hormones, such as catecholamines and cortisol, into the circulation. Catecholamines are released from the chromaffin tissue situated in the head kidney of teleosts and also from the endings of adrenergic nerves. Cortisol is released from the interrenal tissue, located in the head (anterior) kidney, in response to several pituitary hormones, but most potently to adrenocorticotrophic hormone (ACTH). A recent study showed that ACTH may also stimulate adrenaline release and that chronic cortisol treatment may affect catecholamine storage and release in trout. As both the chromaffin tissue and the interrenal tissue lie in close proximity in fish, these results suggest the possibility that a paracrine control for stress hormone regulation exists in fish.
The resting and stressed levels of adrenaline, cortisol, and glucose concentrations in the plasma of salmonids are: adrenaline, < 3 and 20-70 (nmoles/l); cortisol, < 10 and 40-200 (ng/ml); and glucose, <4 and > 5 (mmol/l), respectively. These values should serve as general guidelines since individual conditions, including genetic characteristics, prior rearing history, and local environment, will modify the plasma values for control and stressed states. In addition to the measurement of these stress hormones, the measurement of plasma glucose concentration has been used as an indicator of stressed states in fish. It is probably the most commonly measured secondary change that occurs during the stress response in fish.
Secondary Response
This response comprises the various biochemical and physiological effects associated with stress and is mediated to a large extent by the stress hormones mentioned previously. The stress hormones activate a number of metabolic pathways that result in alterations in blood chemistry and haematology. The section following on the metabolic aspects of stress elaborates on such a secondary response.
Tertiary Response
This response represents whole-animal- and population-level changes associated with stress. If the fish is unable to acclimate or adapt to the stressor, whole-animal changes may occur as a result of energy repartitioning by diverting energy substrates to cope with the enhanced energy demand associated with stress and away from vital life processes such as reproduction and anabolic processes such as growth.
Energy Metabolism and Stress
------------------------------------
Regulation of social behaviour
Oxytocin is a group of neuropeptides found in most vertebrate. One form of oxytocin functions as a hormone which is associated with human love. In 2012, researchers injected cichlids شائكات الزعانف from the social species Neolamprologus pulcher, either with this form of isotocin or with a control saline solution. They found isotocin increased "responsiveness to social information", which suggests "it is a key regulator of social behavior that has evolved and endured since ancient times".
Effects of pollutionFish can bioaccumulate pollutants that are discharged into waterways. Estrogenic compounds found in pesticides, birth control, plastics, plants, fungi, bacteria, and synthetic drugs leeched into rivers are affecting the endocrine systems of native species. In Boulder, Colorado, white sucker fish found downstream of a municipal waste water treatment plant exhibit impaired or abnormal sexual development. The same feminization effect can also be seen in male African clawed frogs when exposed to high levels of atrazine, a widely used pesticide. In the marine ecosystem, organochlorine contaminants like pesticides, herbicides (DDT), and chlordan are accumulating within fish tissue and disrupting their endocrine system. High frequencies of infertility and high levels of organochlorines have been found in bonnethead sharks along the Gulf Coast of Florida. These endocrine-disrupting compounds are similar in structure to naturally occurring hormones in fish. They can modulate hormonal interactions in fish by:
Binding to cellular receptors, causing unpredictable and abnormal cell activity
blocking receptor sites, inhibiting activity promoting the creation of extra receptor sites, amplifying the effects of the hormone or compound interacting with naturally occurring hormones, changing their shape and impact affecting hormone synthesis or metabolism, causing an improper balance or quantity of hormones.
Osmoregulation
Two major types of osmoregulation are osmoconformers and osmoregulators. Osmoconformers match their body osmolarity to their environment actively or passively. Most marine invertebrates are osmoconformers, although their ionic composition may be different from that of seawater.
Osmoregulators tightly regulate their body osmolarity, which always stays constant, and are more common in the animal kingdom. Osmoregulators actively control salt concentrations despite the salt concentrations in the environment. An example is freshwater fish. The gills actively uptake salt from the environment by the use of mitochondria-rich cells. Water will diffuse into the fish, so it excretes a very hypotonic (dilute) urine to expel all the excess water. A marine fish has an internal osmotic concentration lower than that of the surrounding seawater, so it tends to lose water and gain salt. It actively excretes salt out from the gills. Most fish are stenohaline, which means they are restricted to either salt or fresh water and cannot survive in water with a different salt concentration than they are adapted to. However, some fish show a tremendous ability to effectively osmoregulate across a broad range of salinities; fish with this ability are known as euryhaline species, e.g., salmon. Salmon has been observed to inhabit two utterly disparate environments — marine and fresh water — and it is inherent to adapt to both by bringing in behavioral and physiological modifications.
In contrast to bony fish, with the exception of the coelacanth, the blood and other tissue of sharks and Chondrichthyes is generally isotonic to their marine environments because of the high concentration of urea and trimethylamine N-oxide (TMAO), allowing them to be in osmotic balance with the seawater. This adaptation prevents most sharks from surviving in freshwater, and they are therefore confined to marine environments. A few exceptions exist, such as the bull shark, which has developed a way to change its kidney function to excrete large amounts of urea. When a shark dies, the urea is broken down to ammonia by bacteria, causing the dead body to gradually smell strongly of ammonia.
Sharks have adopted a different, efficient mechanism to conserve water, i.e., osmoregulation. They retain urea in their blood in relatively higher concentration. Urea is damaging to living tissue so, to cope لتحمل with this problem, some fish retain trimethylamine oxide. This provides a better solution to urea's toxicity. Sharks, having slightly higher solute concentration (i.e., above 1000 mOsm which is sea solute concentration), do not drink water like fresh water fish.
Pain in fishExperiments done by William Tavolga provide evidence that fish have pain and fear responses. For instance, in Tavolga’s experiments, toadfish grunted when electrically shocked and over time they came to grunt at the mere sight of an electrode.
In 2003, Scottish scientists at the University of Edinburgh and the Roslin Institute concluded that rainbow trout exhibit behaviors often associated with pain as other animals. Bee venom and acetic acid injected into the lips resulted in fish rocking their bodies and rubbing their lips along the sides and floors of their tanks, which the researchers concluded were attempts to relieve pain, similar to what mammals would do. Neurons fired in a pattern resembling human neuronal patterns.
Professor James D. Rose of the University of Wyoming claimed the study was flawed since it did not provide proof that fish possess "conscious awareness, particularly a kind of awareness نوع من الوعي that is meaningfully like ours". Rose argues that since fish brains are so different from human brains, fish are probably not conscious in the manner humans are, so that reactions similar to human reactions to pain instead have other causes. Rose had published a study a year earlier arguing that fish cannot feel pain because their brains lack a neocortex.
Physiological Stress Response
In response to a stressor such as handling or crowding, a fish will undergo a series of biochemical and physiological changes in an attempt to compensate for the challenge imposed upon it and, thereby, cope with the stress. The stress response in fish has been broadly categorized into the primary, secondary, and tertiary responses.
Primary Response
This initial response represents the preception of an altered state and initiates a neuroendocrine/endocrine response that forms part of the generalized stress response in fish. This response includes the rapid release of stress hormones, such as catecholamines and cortisol, into the circulation. Catecholamines are released from the chromaffin tissue situated in the head kidney of teleosts and also from the endings of adrenergic nerves. Cortisol is released from the interrenal tissue, located in the head (anterior) kidney, in response to several pituitary hormones, but most potently to adrenocorticotrophic hormone (ACTH). A recent study showed that ACTH may also stimulate adrenaline release and that chronic cortisol treatment may affect catecholamine storage and release in trout. As both the chromaffin tissue and the interrenal tissue lie in close proximity in fish, these results suggest the possibility that a paracrine control for stress hormone regulation exists in fish.
The resting and stressed levels of adrenaline, cortisol, and glucose concentrations in the plasma of salmonids are: adrenaline, < 3 and 20-70 (nmoles/l); cortisol, < 10 and 40-200 (ng/ml); and glucose, <4 and > 5 (mmol/l), respectively. These values should serve as general guidelines since individual conditions, including genetic characteristics, prior rearing history, and local environment, will modify the plasma values for control and stressed states. In addition to the measurement of these stress hormones, the measurement of plasma glucose concentration has been used as an indicator of stressed states in fish. It is probably the most commonly measured secondary change that occurs during the stress response in fish.
Secondary Response
This response comprises the various biochemical and physiological effects associated with stress and is mediated to a large extent by the stress hormones mentioned previously. The stress hormones activate a number of metabolic pathways that result in alterations in blood chemistry and haematology. The section following on the metabolic aspects of stress elaborates on such a secondary response.
Tertiary Response
This response represents whole-animal- and population-level changes associated with stress. If the fish is unable to acclimate or adapt to the stressor, whole-animal changes may occur as a result of energy repartitioning by diverting energy substrates to cope with the enhanced energy demand associated with stress and away from vital life processes such as reproduction and anabolic processes such as growth.
Energy Metabolism and Stress
- Stress is a energy-demanding process, and the animal has to mobilize energy substrates to metabolically cope with stress. As mentioned above, one of the most common indicators of metabolic effects due to stress is the increase in plasma glucose concentration. The plasma glucose concentration in circulation is dependent on glucose production and its clearance from the circulation. The production of glucose with stress assists the animal by providing energy substrates to tissues such as brain, gills and muscles in order to cope with the increased energy demand. Liver is the main source of glucose production and is achieved by glycogenolysis and/or gluconeogenesis.
- Adrenaline and cortisol have been shown to increase glucose production in fish and may play an important role in the stress-associated increase in plasma glucose concentration. Although adrenaline is cleared rapidly from circulation after a stressor, plasma glucose remains elevated for longer periods of time, suggesting that cortisol (which remains elevated for a longer period of time) plays a role in the long-term maintenance of glucose post-stress in fish.
------------------------------------
Regulation of social behaviour
Oxytocin is a group of neuropeptides found in most vertebrate. One form of oxytocin functions as a hormone which is associated with human love. In 2012, researchers injected cichlids شائكات الزعانف from the social species Neolamprologus pulcher, either with this form of isotocin or with a control saline solution. They found isotocin increased "responsiveness to social information", which suggests "it is a key regulator of social behavior that has evolved and endured since ancient times".
Effects of pollutionFish can bioaccumulate pollutants that are discharged into waterways. Estrogenic compounds found in pesticides, birth control, plastics, plants, fungi, bacteria, and synthetic drugs leeched into rivers are affecting the endocrine systems of native species. In Boulder, Colorado, white sucker fish found downstream of a municipal waste water treatment plant exhibit impaired or abnormal sexual development. The same feminization effect can also be seen in male African clawed frogs when exposed to high levels of atrazine, a widely used pesticide. In the marine ecosystem, organochlorine contaminants like pesticides, herbicides (DDT), and chlordan are accumulating within fish tissue and disrupting their endocrine system. High frequencies of infertility and high levels of organochlorines have been found in bonnethead sharks along the Gulf Coast of Florida. These endocrine-disrupting compounds are similar in structure to naturally occurring hormones in fish. They can modulate hormonal interactions in fish by:
Binding to cellular receptors, causing unpredictable and abnormal cell activity
blocking receptor sites, inhibiting activity promoting the creation of extra receptor sites, amplifying the effects of the hormone or compound interacting with naturally occurring hormones, changing their shape and impact affecting hormone synthesis or metabolism, causing an improper balance or quantity of hormones.
Osmoregulation
Two major types of osmoregulation are osmoconformers and osmoregulators. Osmoconformers match their body osmolarity to their environment actively or passively. Most marine invertebrates are osmoconformers, although their ionic composition may be different from that of seawater.
Osmoregulators tightly regulate their body osmolarity, which always stays constant, and are more common in the animal kingdom. Osmoregulators actively control salt concentrations despite the salt concentrations in the environment. An example is freshwater fish. The gills actively uptake salt from the environment by the use of mitochondria-rich cells. Water will diffuse into the fish, so it excretes a very hypotonic (dilute) urine to expel all the excess water. A marine fish has an internal osmotic concentration lower than that of the surrounding seawater, so it tends to lose water and gain salt. It actively excretes salt out from the gills. Most fish are stenohaline, which means they are restricted to either salt or fresh water and cannot survive in water with a different salt concentration than they are adapted to. However, some fish show a tremendous ability to effectively osmoregulate across a broad range of salinities; fish with this ability are known as euryhaline species, e.g., salmon. Salmon has been observed to inhabit two utterly disparate environments — marine and fresh water — and it is inherent to adapt to both by bringing in behavioral and physiological modifications.
In contrast to bony fish, with the exception of the coelacanth, the blood and other tissue of sharks and Chondrichthyes is generally isotonic to their marine environments because of the high concentration of urea and trimethylamine N-oxide (TMAO), allowing them to be in osmotic balance with the seawater. This adaptation prevents most sharks from surviving in freshwater, and they are therefore confined to marine environments. A few exceptions exist, such as the bull shark, which has developed a way to change its kidney function to excrete large amounts of urea. When a shark dies, the urea is broken down to ammonia by bacteria, causing the dead body to gradually smell strongly of ammonia.
Sharks have adopted a different, efficient mechanism to conserve water, i.e., osmoregulation. They retain urea in their blood in relatively higher concentration. Urea is damaging to living tissue so, to cope لتحمل with this problem, some fish retain trimethylamine oxide. This provides a better solution to urea's toxicity. Sharks, having slightly higher solute concentration (i.e., above 1000 mOsm which is sea solute concentration), do not drink water like fresh water fish.
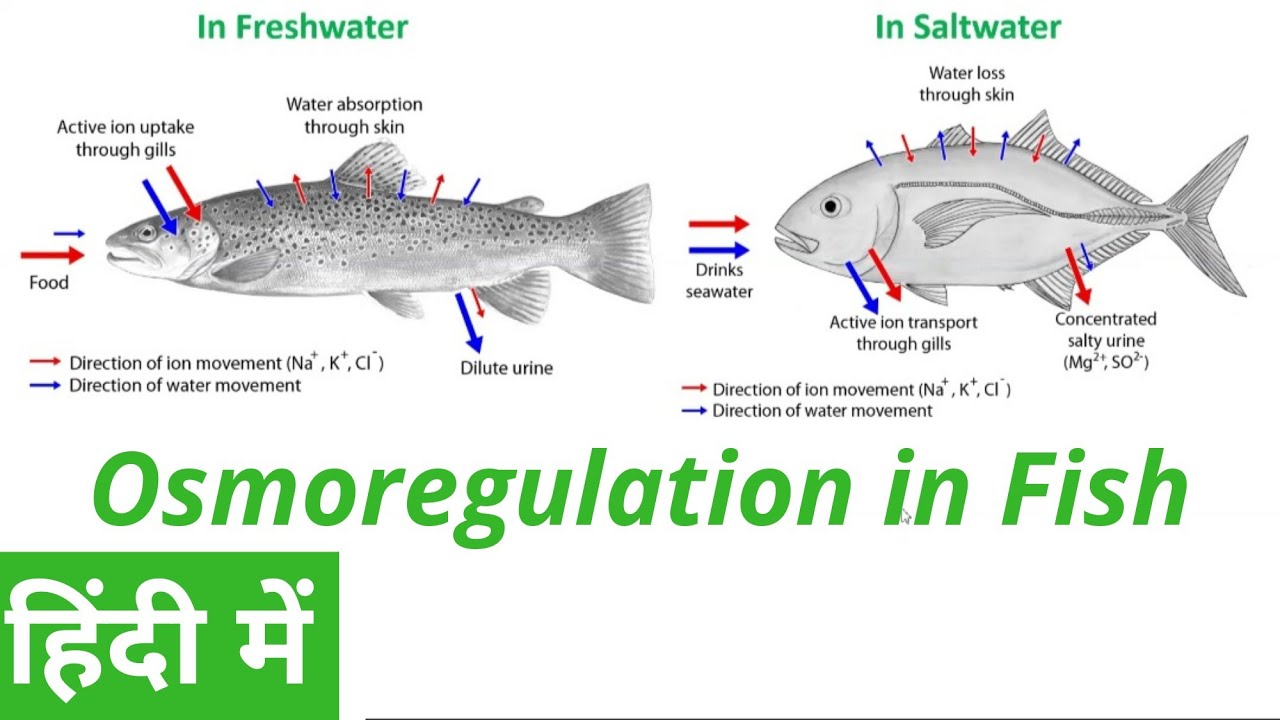
Thermoregulation
Homeothermy and poikilothermy refer to how stable an organism's temperature is. Most endothermic organisms are homeothermic, like mammals. However, animals with facultative endothermy are often poikilothermic, meaning their temperature can vary considerably. Similarly, most fish are ectotherms, as all of their heat comes from the surrounding water. However, most are homeotherms because their temperature is very stable.
Most organisms have a preferred temperature range, however some can be acclimated to temperatures colder or warmer than what they are typically used to. An organism's preferred temperature is typically the temperature at which the organism's physiological processes can act at optimal rates. When fish become acclimated to other temperatures, the efficiency of their physiological processes may decrease but will continue to function. This is called the thermal neutral zone at which an organism can survive indefinitely.
Vernon has done work on the death temperature and paralysis temperature (temperature of heat rigor حرارة صارمة) of various animals. He found that species of the same class showed very similar temperature values, those from the Amphibia examined being 38.5 °C, fish 39 °C, Reptilia 45 °C, and various Molluscs 46 °C.
To cope with low temperatures, some fish have developed the ability to remain functional even when the water temperature is below freezing; some use natural antifreeze or antifreeze proteins to resist ice crystal formation in their tissues.
Most sharks are "cold-blooded" or, more precisely, poikilothermic, meaning that their internal body temperature matches that of their ambient environment. Members of the family Lamnidae (such as the shortfin mako shark and the great white shark) are homeothermic and maintain a higher body temperature than the surrounding water. In these sharks, a strip of aerobic red muscle located near the center of the body generates the heat, which the body retains via a countercurrent exchange mechanism by a system of blood vessels called the rete mirabile ("miraculous net"). The common thresher shark has a similar mechanism for maintaining an elevated body temperature, which is thought to have evolved independently.
Tuna can maintain the temperature of certain parts of their body above the temperature of ambient seawater. For example, blue fin tuna maintain a core body temperature of 25–33 °C (77–91 °F), in water as cold as 6 °C (43 °F). However, unlike typical endothermic creatures such as mammals and birds, tuna do not maintain temperature within a relatively narrow range. Tuna achieve endothermy by conserving the heat generated through normal metabolism. The rete mirabile ("wonderful net"), the intertwining of veins and arteries in the body's periphery, transfers heat from venous blood to arterial blood via a counter-current exchange system, thus mitigating the effects of surface cooling. This allows the tuna to elevate the temperatures of the highly aerobic tissues of the skeletal muscles, eyes and brain, which supports faster swimming speeds and reduced energy expenditure, and which enables them to survive in cooler waters over a wider range of ocean environments than those of other fish. In all tunas, however, the heart operates at ambient temperature, as it receives cooled blood, and coronary circulation is directly from the gills.
Homeothermy: Although most fish are exclusively ectothermic, there are exceptions. Certain species of fish maintain elevated body temperatures. Endothermic teleosts (bony fish) are all in the suborder Scombroidei and include the billfishes, tunas, including a "primitive" mackerel species, Gasterochisma melampus. All sharks in the family Lamnidae – shortfin mako, long fin mako, white, porbeagle, and salmon shark – are endothermic, and evidence suggests the trait exists in family Alopiidae (thresher sharks). The degree of endothermy varies from the billfish, which warm only their eyes and brain, to bluefin tuna and porbeagle sharks who maintain body temperatures elevated in excess of 20 °C above ambient water temperatures.[38] See also gigantothermy. Endothermy, though metabolically costly, is thought to provide advantages such as increased muscle strength, higher rates of central nervous system processing, and higher rates of digestion.
In some fish, a rete mirabile allows for an increase in muscle temperature in regions where this network of vein and arteries is found. The fish is able to thermoregulate certain areas of their body. Additionally, this increase in temperature leads to an increase in basal metabolic temperature. The fish is now able to split ATP at a higher rate and ultimately can swim faster.
The eye of a swordfish can generate heat to better cope with detecting their prey at depths of 2000 feet.
-------------------------------------------------------------------------------
Muscular System
Fish swim by contracting longitudinal red muscle and obliquely oriented white muscles. The red muscle is aerobic and needs oxygen which is supplied by myoglobin. The white muscle is anaerobic and it does not need oxygen. Red muscles are used for sustained activity such as cruising at slow speeds on ocean migrations. White muscles are used for bursts of activity, such as jumping or sudden bursts of speed for catching prey.
Mostly fish have white muscles, but the muscles of some fishes, such as scombroids and salmonids, range from pink to dark red. The red myotomal muscles derive their colour from myoglobin, an oxygen-binding molecule, which tuna express in quantities far higher than most other fish. The oxygen-rich blood further enables energy delivery to their muscles.
Most fish move by alternately contracting paired sets of muscles on either side of the backbone. These contractions form S-shaped curves that move down the body. As each curve reaches the back fin, backward force is applied to the water, and in conjunction with the fins, moves the fish forward. The fish's fins function like an airplane's flaps. Fins also increase the tail's surface area, increasing speed. The streamlined body of the fish decreases the amount of friction from the water.
A typical characteristic of many animals that utilize undulatory locomotion is that they have segmented muscles, or blocks of myomeres, running from their head to tails which are separated by connective tissue called myosepta. In addition, some segmented muscle groups, such the lateral hypaxial musculature in the salamander are oriented at an angle to the longitudinal direction. For these obliquely oriented fibers the strain in the longitudinal direction is greater than the strain in the muscle fiber direction leading to an architectural gear ratio greater than 1. A higher initial angle of orientation and more dorsoventral bulging produces a faster muscle contraction but results in a lower amount of force production. It is hypothesized that animals employ a variable gearing mechanism that allows self-regulation of force and velocity to meet the mechanical demands of the contraction. When a pennate muscle is subjected to a low force, resistance to width changes in the muscle cause it to rotate which consequently produce a higher architectural gear ratio (AGR) (high velocity). However, when subject to a high force, the perpendicular fiber force component overcomes the resistance to width changes and the muscle compresses producing a lower AGR (capable of maintaining a higher force output).
Homeothermy and poikilothermy refer to how stable an organism's temperature is. Most endothermic organisms are homeothermic, like mammals. However, animals with facultative endothermy are often poikilothermic, meaning their temperature can vary considerably. Similarly, most fish are ectotherms, as all of their heat comes from the surrounding water. However, most are homeotherms because their temperature is very stable.
Most organisms have a preferred temperature range, however some can be acclimated to temperatures colder or warmer than what they are typically used to. An organism's preferred temperature is typically the temperature at which the organism's physiological processes can act at optimal rates. When fish become acclimated to other temperatures, the efficiency of their physiological processes may decrease but will continue to function. This is called the thermal neutral zone at which an organism can survive indefinitely.
Vernon has done work on the death temperature and paralysis temperature (temperature of heat rigor حرارة صارمة) of various animals. He found that species of the same class showed very similar temperature values, those from the Amphibia examined being 38.5 °C, fish 39 °C, Reptilia 45 °C, and various Molluscs 46 °C.
To cope with low temperatures, some fish have developed the ability to remain functional even when the water temperature is below freezing; some use natural antifreeze or antifreeze proteins to resist ice crystal formation in their tissues.
Most sharks are "cold-blooded" or, more precisely, poikilothermic, meaning that their internal body temperature matches that of their ambient environment. Members of the family Lamnidae (such as the shortfin mako shark and the great white shark) are homeothermic and maintain a higher body temperature than the surrounding water. In these sharks, a strip of aerobic red muscle located near the center of the body generates the heat, which the body retains via a countercurrent exchange mechanism by a system of blood vessels called the rete mirabile ("miraculous net"). The common thresher shark has a similar mechanism for maintaining an elevated body temperature, which is thought to have evolved independently.
Tuna can maintain the temperature of certain parts of their body above the temperature of ambient seawater. For example, blue fin tuna maintain a core body temperature of 25–33 °C (77–91 °F), in water as cold as 6 °C (43 °F). However, unlike typical endothermic creatures such as mammals and birds, tuna do not maintain temperature within a relatively narrow range. Tuna achieve endothermy by conserving the heat generated through normal metabolism. The rete mirabile ("wonderful net"), the intertwining of veins and arteries in the body's periphery, transfers heat from venous blood to arterial blood via a counter-current exchange system, thus mitigating the effects of surface cooling. This allows the tuna to elevate the temperatures of the highly aerobic tissues of the skeletal muscles, eyes and brain, which supports faster swimming speeds and reduced energy expenditure, and which enables them to survive in cooler waters over a wider range of ocean environments than those of other fish. In all tunas, however, the heart operates at ambient temperature, as it receives cooled blood, and coronary circulation is directly from the gills.
Homeothermy: Although most fish are exclusively ectothermic, there are exceptions. Certain species of fish maintain elevated body temperatures. Endothermic teleosts (bony fish) are all in the suborder Scombroidei and include the billfishes, tunas, including a "primitive" mackerel species, Gasterochisma melampus. All sharks in the family Lamnidae – shortfin mako, long fin mako, white, porbeagle, and salmon shark – are endothermic, and evidence suggests the trait exists in family Alopiidae (thresher sharks). The degree of endothermy varies from the billfish, which warm only their eyes and brain, to bluefin tuna and porbeagle sharks who maintain body temperatures elevated in excess of 20 °C above ambient water temperatures.[38] See also gigantothermy. Endothermy, though metabolically costly, is thought to provide advantages such as increased muscle strength, higher rates of central nervous system processing, and higher rates of digestion.
In some fish, a rete mirabile allows for an increase in muscle temperature in regions where this network of vein and arteries is found. The fish is able to thermoregulate certain areas of their body. Additionally, this increase in temperature leads to an increase in basal metabolic temperature. The fish is now able to split ATP at a higher rate and ultimately can swim faster.
The eye of a swordfish can generate heat to better cope with detecting their prey at depths of 2000 feet.
-------------------------------------------------------------------------------
Muscular System
Fish swim by contracting longitudinal red muscle and obliquely oriented white muscles. The red muscle is aerobic and needs oxygen which is supplied by myoglobin. The white muscle is anaerobic and it does not need oxygen. Red muscles are used for sustained activity such as cruising at slow speeds on ocean migrations. White muscles are used for bursts of activity, such as jumping or sudden bursts of speed for catching prey.
Mostly fish have white muscles, but the muscles of some fishes, such as scombroids and salmonids, range from pink to dark red. The red myotomal muscles derive their colour from myoglobin, an oxygen-binding molecule, which tuna express in quantities far higher than most other fish. The oxygen-rich blood further enables energy delivery to their muscles.
Most fish move by alternately contracting paired sets of muscles on either side of the backbone. These contractions form S-shaped curves that move down the body. As each curve reaches the back fin, backward force is applied to the water, and in conjunction with the fins, moves the fish forward. The fish's fins function like an airplane's flaps. Fins also increase the tail's surface area, increasing speed. The streamlined body of the fish decreases the amount of friction from the water.
A typical characteristic of many animals that utilize undulatory locomotion is that they have segmented muscles, or blocks of myomeres, running from their head to tails which are separated by connective tissue called myosepta. In addition, some segmented muscle groups, such the lateral hypaxial musculature in the salamander are oriented at an angle to the longitudinal direction. For these obliquely oriented fibers the strain in the longitudinal direction is greater than the strain in the muscle fiber direction leading to an architectural gear ratio greater than 1. A higher initial angle of orientation and more dorsoventral bulging produces a faster muscle contraction but results in a lower amount of force production. It is hypothesized that animals employ a variable gearing mechanism that allows self-regulation of force and velocity to meet the mechanical demands of the contraction. When a pennate muscle is subjected to a low force, resistance to width changes in the muscle cause it to rotate which consequently produce a higher architectural gear ratio (AGR) (high velocity). However, when subject to a high force, the perpendicular fiber force component overcomes the resistance to width changes and the muscle compresses producing a lower AGR (capable of maintaining a higher force output).
Most fishes bend as a simple, homogenous beam during swimming via contractions of longitudinal red muscle fibers and obliquely oriented white muscle fibers within the segmented axial musculature. The fiber strain (εf) experienced by the longitudinal red muscle fibers is equivalent to the longitudinal strain (εx). The deeper white muscle fibers fishes show diversity in arrangement. These fibers are organized into cone-shaped structures and attach to connective tissue sheets known as myosepta; each fiber shows a characteristic dorsoventral (α) and mediolateral (φ) trajectory. The segmented architecture theory predicts that, εx > εf. This phenomenon results in an architectural gear ratio, determined as longitudinal strain divided by fiber strain (εx / εf), greater than one and longitudinal velocity amplification; furthermore, this emergent velocity amplification may be augmented by variable architectural gearing via mesolateral and dorsoventral shape changes, a pattern seen in pennate muscle contractions. A red-to-white gearing ratio (red εf / white εf) captures the combined effect of the longitudinal red muscle fiber and oblique white muscle fiber strains.

Buoyancy
The body of a fish is denser than water, so fish must compensate for the difference or they will sink. Many bony fishes have an internal organ called a swim bladder, or gas bladder, that adjusts their buoyancy through manipulation of gases. In this way, fish can stay at the current water depth, or ascend or descend without having to waste energy in swimming. The bladder is only found in bony fishes. In the more primitive groups like some minnows, bichirs and lungfish, the bladder is open to the esophagus and double as a lung. It is often absent in fast swimming fishes such as the tuna and mackerel families. The condition of a bladder open to the esophagus is called physostome, the closed condition physoclist. In the latter, the gas content of the bladder is controlled through the rete mirabilis, a network of blood vessels effecting gas exchange between the bladder and the blood.
In some fish, a rete mirabile fills the swim bladder with oxygen. A countercurrent exchange system is utilized between the venous and arterial capillaries. By lowering the pH levels in the venous capillaries, oxygen unbinds from blood hemoglobin. This causes an increase in venous blood oxygen concentration, allowing the oxygen to diffuse through the capillary membrane and into the arterial capillaries, where oxygen is still sequestered to hemoglobin. The cycle of diffusion continues until the concentration of oxygen in the arterial capillaries is supersaturated(larger than the concentration of oxygen in the swim bladder). At this point, the free oxygen in the arterial capillaries diffuses into the swim bladder via the gas gland.
Unlike bony fish, sharks do not have gas-filled swim bladders for buoyancy. Instead, sharks rely on a large liver filled with oil that contains squalene, and their cartilage, which is about half the normal density of bone. Their liver constitutes up to 30% of their total body mass. The liver's effectiveness is limited, so sharks employ dynamic lift to maintain depth when not swimming. Sand tiger sharks store air in their stomachs, using it as a form of swim bladder. Most sharks need to constantly swim in order to breathe and cannot sleep very long without sinking (if at all). However, certain species, like the nurse shark, are capable of pumping water across their gills, allowing them to rest on the ocean bottom. Sharks, like this three tonne great white shark, don't have swim bladders. Most sharks need to keep swimming to avoid sinking.

References
Andrews, Chris; Adrian Exell; Neville Carrington (2003): Manual Of Fish Health. Firefly Books.
Arukwe, Augustine (2001): "Cellular and Molecular Responses to Endocrine-Modulators and the Impact on Fish Reproduction". Marine Pollution Bulletin. 42 (8): 643.
Arukwe, Augustine (August 2001). "Cellular and Molecular Responses to Endocrine-Modulators and The Impact on Fish Reproduction". Marine Pollution Bulletin. 42 (8): 643. doi:10.1016/S0025-326X(01)00062-5.
Clack, J. A. (2002): Gaining ground: the origin and evolution of tetrapods. Indiana University Press, Bloomington, Indiana. 369 pp
Compagno, Leonard; Dando, Marc; Fowler, Sarah (2005): Sharks of the World. Collins Field Guides. ISBN 0-00-713610-2. OCLC 183136093.
Fowler, Sarah; Cavanagh, Rachel.(2017): "Sharks, Rays, and Chimaeras: The Status of Chondrichthyan Fishes" (PDF). The World Conservation Union. Retrieved 3 May 2017.
Gilbertson, Lance (1999): Zoology Laboratory Manual. New York: McGraw-Hill Companies, Inc. ISBN 0-07-237716-.
Guillaume, Jean; Praxis Publishing; Sadasivam Kaushik; Pierre Bergot; Robert Metailler (2001): Nutrition and Feeding of Fish and Crustaceans. Springer. p. 31. ISBN 978-1-85233-241-9. Retrieved 2009-01-09.
Hayes, Tyron; Narayan, Anne (January 2010): "Atrazine induces complete feminization and chemical castration in male African clawed frogs (Xenopus laevis)". Proceedings of the National Academy of Sciences. 107 (10): 4612.
Hoar WS and Randall DJ (1984): Fish Physiology: Gills: Part B – Ion and water transferAcademic Press. ISBN 978-0-08-058532-1.
John A. Musick (2005): "Management techniques for elasmobranch fisheries: 14. Shark Utilization". FAO: Fisheries and Aquaculture Department. Retrieved 2008-03-16.
Laurin M. (1998): The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I-systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: pp 1-42.
Martin, R. Aidan (April 1992): "Fire in the Belly of the Beast". ReefQuest Centre for Shark Research. Retrieved 2009-08-21.
Pinto, Patricia; Estevao, Maria; Power, Deborah (August 2014): "Effects of Estrogens and Estrogenic Disrupting Compounds on Fish Mineralized Tissues". Marine Drugs. 12 (8): 4474. doi:10.3390/md12084474.
Prosser, C. Ladd (1991): Comparative Animal Physiology, Environmental and Metabolic Animal Physiology (4th ed.). Hoboken, NJ: Wiley-Liss. pp. 1–12. ISBN 0-471-85767.
Reddon, AR; O'Connor, CM; Marsh-Rollo, SE; Balcshine, S (2012): "Effects of isotocin on social responses in a cooperatively breeding fish". Animal Behaviour. 84 (4): 753–760. doi:10.1016/j.anbehav.2012.07.021.
Romer, Alfred Sherwood; Parsons, Thomas S. (1977):The Vertebrate Body. Philadelphia, PA: Holt-Saunders International. pp. 316–327. ISBN 0-03-910284-.
Schwab, Ivan R (2002): "More than just cool shades". British Journal of Ophthalmology. 86: 1075. doi:10.1136/bjo.86.10.1075.
Scott, Thomas (1996): Concise encyclopedia biology. Walter de Gruyter. p. 542. ISBN 978-3-11-010661-9.
Sepulveda, C.A.; Dickson, K.A.; Bernal, D.; Graham, J. B. (2008): "Elevated red myotomal muscle temperatures in the most basal tuna species, Allothunnus fallai"(PDF). Journal of Fish Biology. 73 (1): 241–249.
Thomas Batten (2008): "MAKO SHARK Isurus oxyrinchus". Delaware Sea Grant, University of Delaware. Retrieved 2008-03-16.
Wilmer, Pat; Stone, Graham; Johnston, Ian (2009): Environmental Physiology of Animals. Wiley. pp. 198–200. ISBN 978-1-4051-0724-2.
